14. Data Processing, Evaluation, and Analysis
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
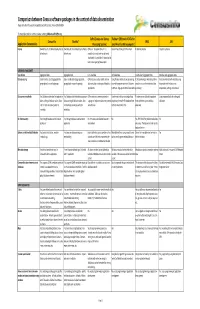
Comparison Between Census Software Packages in the Context of Data Dissemination Prepared by Devinfo Support Group (DSG) and UN Statistics Division (DESA/UNSD)
Comparison between Census software packages in the context of data dissemination Prepared by DevInfo Support Group (DSG) and UN Statistics Division (DESA/UNSD) For more information or comments please contact: [email protected] CsPro (Census and Survey Redatam (REtrieval of DATa for CensusInfo DevInfo* SPSS SAS Application Characteristics Processing System) small Areas by Microcomputer) License Owned by UN, distributed royalty-free to Owned by UN, distributed royalty-free to CSPro is in the public domain. It is Version free of charge for Download Properitery license Properitery license all end-users all end-users available at no cost and may be freely distributed. It is available for download at www.census.gov/ipc/www/cspro DATABASE MANAGEMENT Type of data Aggregated data. Aggregated data. Individual data. Individual data. Individual and aggregated data. Individual and aggregated data. Data processing Easily handles data disaggregated by Easily handles data disaggregated by CSPro lets you create, modify, and run Using Process module allows processing It allows entering primary data, define It lets you interact with your data using geographical area and subgroups. geographical area and subgroups. data entry, batch editing, and tabulation data with programs written in Redatam variables and perform statistical data integrated tools for data entry, applications command language or limited Assisstants processing computation, editing, and retrieval. Data consistency checks Yes, Database administration application Yes, Database administration -

Research; *Electronic Data Processing; Information Processing; Information Systems; *Manpower Utilization; *Personnel Data; Personnel Management; Questionnaires
DOCUMENT RESUME ED 084 382 CE 000 533 AUTHOR Bayhylle, J. E., Ed.; Hersleb, A., Ed. TITLE The Development of Electronic Data Processing in Manpower Areas; A Comparative Study of Personnel Information Systems and their Implications in Six European Countries. INSTITUTION Organisation for Economic Cooperation and Development, Paris (France) . PUB DATE 73 NOTE 366p. AVAILABLE FROMOECD Publications Center, Suite 1207, 1750 Pennsylvania Ave., N.W., Washington, DC 20006 ($8.50) EDRS PRICE MF-$0.65 HC- $13.16 DESCRIPTORS Case Studies; Data Bases; Data Processing; *Ecqnomic .Research; *Electronic Data Processing; Information Processing; Information Systems; *Manpower Utilization; *Personnel Data; Personnel Management; Questionnaires ABSTRACT Usable data about human resources in the ecqnqmy are fundamentally important, but present methods of obtaining, organizing, and using such information are increasingly unsatisfactory. The application of electronic data processing to manpower and social concerns should be improved and increased. This could stimulate closer exchange between government, institutions, and business enterprises in the collection and dissemination of manpower data. The first phase of the study was a broad-scope questionnaire of 38 companies in Austria, France, Germany, Great Britain, Sweden, and Switzerland which have developed, or are planning to develop, computer-based personnel information, systems. The second phase consisted of depth study of eight of the companies. Case study investigations were concerned with the content and function of the system and with the administrative and personnel consequences of the introduction of such systems. A set of key.points which emerged from the study is developed. A model demonstrates the need for coordination to increase communication vertically and horizontally. -

Applied Accounting and Automatic Data-Processing Systems." (1962)
Louisiana State University LSU Digital Commons LSU Historical Dissertations and Theses Graduate School 1962 Applied Accounting and Automatic Data- Processing Systems. William Elwood Swyers Louisiana State University and Agricultural & Mechanical College Follow this and additional works at: https://digitalcommons.lsu.edu/gradschool_disstheses Recommended Citation Swyers, William Elwood, "Applied Accounting and Automatic Data-Processing Systems." (1962). LSU Historical Dissertations and Theses. 745. https://digitalcommons.lsu.edu/gradschool_disstheses/745 This Dissertation is brought to you for free and open access by the Graduate School at LSU Digital Commons. It has been accepted for inclusion in LSU Historical Dissertations and Theses by an authorized administrator of LSU Digital Commons. For more information, please contact [email protected]. This dissertation has been 62—3671 microfilmed exactly as received SWYERS, William Elwood, 1922- APPLIED ACCOUNTING AND AUTOMATIC DATA-PRO CESSING SYSTEMS. Louisiana State University, Ph.D., 1962 Economics, commerce—business University Microfilms, Inc., Ann Arbor, Michigan APPLIED ACCOUNTING AND AUTOMATIC DATA-PROCESSING SYSTEMS A Dissertation Submitted to the Graduate Faculty of the Louisiana State University and Agricultural and Mechanical College in partial fulfillment of the requirements for the degree of Doctor of Philosophy in The Department of Accounting by William Eiu<Swyers B.S., Centenary College, 1942 M.B.A. t Louisiana State University, 1948 January, 1962 ACKNOWLEDGMENTS The writer wishes to express appreciation to Dr. Robert H. Van Voorhis, Professor of Accounting, Louisiana State University, for his valuable assistance and guidance in the preparation of this dissertation. The writer wishes to acknowledge, also, the helpful suggestions made by Dr. W. D. Ross, Dean of the College of Business Administration, Dr. -

STANDARDS and GUIDELINES for STATISTICAL SURVEYS September 2006
OFFICE OF MANAGEMENT AND BUDGET STANDARDS AND GUIDELINES FOR STATISTICAL SURVEYS September 2006 Table of Contents LIST OF STANDARDS FOR STATISTICAL SURVEYS ....................................................... i INTRODUCTION......................................................................................................................... 1 SECTION 1 DEVELOPMENT OF CONCEPTS, METHODS, AND DESIGN .................. 5 Section 1.1 Survey Planning..................................................................................................... 5 Section 1.2 Survey Design........................................................................................................ 7 Section 1.3 Survey Response Rates.......................................................................................... 8 Section 1.4 Pretesting Survey Systems..................................................................................... 9 SECTION 2 COLLECTION OF DATA................................................................................... 9 Section 2.1 Developing Sampling Frames................................................................................ 9 Section 2.2 Required Notifications to Potential Survey Respondents.................................... 10 Section 2.3 Data Collection Methodology.............................................................................. 11 SECTION 3 PROCESSING AND EDITING OF DATA...................................................... 13 Section 3.1 Data Editing ........................................................................................................ -

What Can a Computer Do? a Computer Is an Electronic Device
You have learnt various methods of data processing and representation that you can use to analyse the geographical phenomena in the preceding chapters. You must have observed that these methods are time consuming and tedious. Have you ever thought of a method of data processing and their graphical representation that can save time and improve efficiency? If you have used a computer for word processing, then you must have noticed that the computer is more versatile as it facilitates the onscreen editing of the text, copy and move it from one place to another, or even delete the unwanted text. Similarly, the computer may also be used for data processing, preparation of diagrams/graphs and drawing of maps, provided you have an access to the related application software. In other words, a computer can be used for a wide range of applications. It must, however, be clearly understood that a computer carries out the instructions it receives from the users. In other words, it cannot perform any function on its own. In the present chapter, we will discuss the use of computers in data processing and mapping. What can a Computer do? A computer is an electronic device. It consists of various sub-systems, like memory, micro-processor, input system and output system. All these sub- systems work together to make it an integrated system. It is an extremely powerful device, which is apt to have an important effect on the systems of data processing, mapping and analysis. A computer is a fast and a versatile machine that can perform simple arithmetic operations, such as, addition, subtraction, multiplication and division, and can also solve complex mathematical, formulae. -

Course Title: Fundermentals of Data Processing
NATIONAL OPEN UNIVERSITY OF NIGERIA SCHOOL OF EDUCATION COURSE CODE: BED 212: COURSE TITLE: FUNDERMENTALS OF DATA PROCESSING BED 212: FUNDERMENTALS OF DATA PROCESSING COURSE DEVELOPER & WRITER: DR. OPATEYE JOHNSON AYODELE School of Education National Open University of Nigeria 14/16 Ahmadu Bello Way, Victoria Island Lagos. Nigeria. COURSE CORDINATOR: DR. INEGBEDION, JULIET O. School of Education National Open University of Nigeria 14/16 Ahmadu Bello Way, Victoria Island Lagos. Nigeria. INTRODUCTION BED 212: FUNDERMENTALS OF DATA PROCESSING This course is designed to equip the students with knowledge of the data processing in technical and vocational education (TVE) especially Business Education research. WHAT YOU WILL LEARN You will learn the components of data processing, hardware and software components, file management and organization in Business Education- based data. It will also interest you to learn about basics of research, approaches and designs with data gathering techniques. Relevant statistical tools you can use to analyse your data would be across and how to write your research reports. COURSE AIMS This course aims at producing competent business educators who will be versed in organizational and research data processing in order to foster time and effective information that will be used in decision making. In order to enable you meet the above aims, modules constituting of units have been produced for your study. Apart from meeting the aims of the course as a whole, each course unit consists of learning objectives which are intended to ensure your learning effectiveness. COURSE OBJECTIVES The course objectives are meant to enable you achieve/acquire the following: 1) Gain in-depth knowledge of data processing and its functions in business organisations and educational settings. -

Hyperspectral Data Processing. Algorithm Design and Analysis
Brochure More information from http://www.researchandmarkets.com/reports/2293144/ Hyperspectral Data Processing. Algorithm Design and Analysis Description: Hyperspectral Data Processing: Algorithm Design and Analysis is a culmination of the research conducted in the Remote Sensing Signal and Image Processing Laboratory (RSSIPL) at the University of Maryland, Baltimore County. Specifically, it treats hyperspectral image processing and hyperspectral signal processing as separate subjects in two different categories. Most materials covered in this book can be used in conjunction with the author’s first book, Hyperspectral Imaging: Techniques for Spectral Detection and Classification, without much overlap. Many results in this book are either new or have not been explored, presented, or published in the public domain. These include various aspects of endmember extraction, unsupervised linear spectral mixture analysis, hyperspectral information compression, hyperspectral signal coding and characterization, as well as applications to conceal target detection, multispectral imaging, and magnetic resonance imaging. Hyperspectral Data Processing contains eight major sections: - Part I: provides fundamentals of hyperspectral data processing - Part II: offers various algorithm designs for endmember extraction - Part III: derives theory for supervised linear spectral mixture analysis - Part IV: designs unsupervised methods for hyperspectral image analysis - Part V: explores new concepts on hyperspectral information compression - Parts VI & VII: develops -

Cit 213 Course Title: Elementary Data Processing
NATIONAL OP EN UNIVERSITY OF NIGERIA SCHOOL OF SCIENCE AND TECHNOLOGY COURSE CODE: CIT 213 COURSE TITLE: ELEMENTARY DATA PROCESSING CO URSE GUIDE CIT 213 ELEMENTARY DATA PROCESSING Course Developer Dr. Ikhu-Omoregbe N. A. Course Editor Course Co-ordinator Afolorunso, A. A. National Open University of Nigeria Lagos. NATIONAL OPEN UNIVERSITY OF NIGERIA National Open University of Nigeria Headquarters 14/16 Ahmadu Bello Way Victoria Island Lagos Abuja Annex 245 Samuel Adesujo Ademulegun Street Central Business District Opposite Arewa Suites Abuja e-mail: [email protected] URL: www.nou.edu.ng National Open University of Nigeria 2008 First Printed 2008 ISBN All Rights Reserved Printed by .. For National Open University of Nigeria TABLE OF CONTENTS PAGE Introduction.............................................................................. 1 - 2 What you will learn in this Course..................................... 2 Course Aims............................................................................... 2 Course Objectives...................................................................... 2 3 Working through this Course.................................................... 3 Course Materials...........................................................................3 Study Units ............................................................................... 3 - 4 Textbooks and References ........................................................ 4 - 5 Assignment File....................................................................... -

Data Processing Operations Technician
Fremont Unified School District Classified Job Description Office and Technical DATA PROCESSING OPERATIONS TECHNICIAN DEFINITION Under general supervision, assists in the day-to-day operation of the Management Information Systems Department; assists users with centralized MIS and/or help desk requests; operates electronic computer and peripheral equipment; and performs related work as assigned. CLASS CHARACTERISTICS This class provides a variety of technical assistance to the Management Information Systems Department, in the areas of tracking and processing mainframe operations jobs and in providing technical assistance to users in various District offices and school sites. This class is distinguished from Computer Operator/Programmer in that the latter performs programming activities in addition to batch and on-line operations within the MIS centralized operations center. EXAMPLES OF DUTIES Receives users requests; determines the appropriate staff to fulfill the request and processes required paperwork; refers special requests to programmers or other technical staff. Schedules batch jobs, ensures and tracks proper sequence to meet established deadlines and user needs. Operates the mainframe console and peripheral equipment to run batch jobs and/or initiates command files for later execution. Obtains printouts of reports to be distributed. Loads appropriate forms and paper styles on line printers. Prepares tapes for microfiche or other long-term storage; calls for pick-up, files and distributes when returned. Maintains database elements, ensuring continuing validity. Assists Operations Supervisor in evaluating operation of the computer and related equipment and in maintaining the system communication scheme. Provides information and assistance to system-users, facilitating on-line applications. In event of malfunction, resets the current operation or contacts appropriate District or vendor support staff as required. -

An Analysis of Information Technology on Data Processing by Using Cobit Framework
(IJACSA) International Journal of Advanced Computer Science and Applications, Vol. 6, No. 9, 2015 An Analysis of Information Technology on Data Processing by using Cobit Framework Surni Erniwati Nina Kurnia Hikmawati Academy of Information Management and Computer Telkom University Bandung AMIKOM Mataram Universitas Telkom Bandung Mataram, Indonesia Bandung, Indonesian Abstract—Information technology and processes is inter administrative services and academic information quickly and connected, directing and controlling the company in achieving accurately. ABASA already supported by IT in data corporate goals through value-added and balancing the risks and processing, which for the procurement of IT is implemented by benefits of information technology. This study is aimed to analyze a separate division, namely division BPSI. However, research the level of maturity (maturity level) on the data process and has not been done to measure the maturity level data produced information technology recommendations that can be processing and mapping the information technology process made as regards the management of IT to support the academic maturity level of the institution cannot be measured, in addition performance of the service to be better. Maturity level to these problems relating to data processing, how to maintain calculation was done by analyzing questionnaires on the state of the completeness, accuracy and availability of data through the information technology. The results of this study obtainable that efforts of data backup, data storage or deletion of data. Based the governance of information technology in data processing in Mataram ASM currently quite good. Current maturity value for on those problems, this study is aimed to make a the data processing has the value 2.69. -

Processing the Data
CHAPTER 7 PROCESSING THE DATA This chapter is written for survey coordinators, data processing experts and technical resource persons. It provides information on how to: Prepare for processing the data Set up a system for managing data processing Carry out data entry Edit the data and create a ‘clean’ data file for analysis Produce tabulations with the indicators Archive and distribute data The MICS3 data-processing system is designed to deliver the first results of a survey within a few weeks after the end of fieldwork. This chapter contains information that will help you to undertake the planning and advance preparation that will make this goal a reality. The chapter begins by giving you an overview of the MICS3 data-processing system. It then discusses each of its components in detail, providing references to supplemental sources of information where appropriate. It closes with a set of three checklists that will help you make the processing of your survey data a success. OVERVIEW The reason that the MICS3 data-processing system can achieve such rapid turnaround time is because data is processed in tandem with survey fieldwork. Data for each cluster is stored in a separate data file and is processed as soon as the questionnaires are returned from the field. This approach breaks data processing down into discrete segments and allows it to progress while fieldwork is ongoing. Thus, by the time the last questionnaires are finished and returned to headquarters, most of the data have already been processed. Processing the data by clusters is not difficult, but it does require meticulous organization. -

A Handbook of Statistical Analyses Using SPSS
A Handbook of Statistical Analyses using SPSS Sabine Landau and Brian S. Everitt CHAPMAN & HALL/CRC A CRC Press Company Boca Raton London New York Washington, D.C. © 2004 by Chapman & Hall/CRC Press LLC Library of Congress Cataloging-in-Publication Data Landau, Sabine. A handbook of statistical analyses using SPSS / Sabine, Landau, Brian S. Everitt. p. cm. Includes bibliographical references and index. ISBN 1-58488-369-3 (alk. paper) 1. SPSS ( Computer file). 2. Social sciences—Statistical methods—Computer programs. 3. Social sciences—Statistical methods—Data processing. I. Everitt, Brian S. II. Title. HA32.E93 2003 519.5d0285—dc22 2003058474 This book contains information obtained from authentic and highly regarded sources. Reprinted material is quoted with permission, and sources are indicated. A wide variety of references are listed. Reasonable efforts have been made to publish reliable data and information, but the author and the publisher cannot assume responsibility for the validity of all materials or for the consequences of their use. Neither this book nor any part may be reproduced or transmitted in any form or by any means, electronic or mechanical, including photocopying, microfilming, and recording, or by any information storage or retrieval system, without prior permission in writing from the publisher. The consent of CRC Press LLC does not extend to copying for general distribution, for promotion, for creating new works, or for resale. Specific permission must be obtained in writing from CRC Press LLC for such copying. Direct all inquiries to CRC Press LLC, 2000 N.W. Corporate Blvd., Boca Raton, Florida 33431. Trademark Notice: Product or corporate names may be trademarks or registered trademarks, and are used only for identification and explanation, without intent to infringe.