Anatomy of the Human Mammary Gland: Current Status of Knowledge
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
Dr. Hale's Lactation Risk Categories
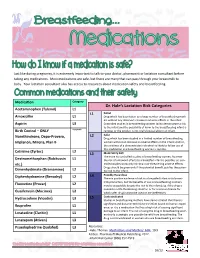
Just like during pregnancy, it is extremely important to talk to your doctor, pharmacist or lactation consultant before taking any medications. Most medications are safe, but there are many that can pass through your breastmilk to baby. Your lactation consultant also has access to resources about medication safety and breastfeeding. Medication Category Dr. Hale’s Lactation Risk Categories Acetaminophen (Tylenol) L1 L1 Safest Amoxicillin L1 Drug which has been taken by a large number of breastfeeding moth- ers without any observed increase in adverse effects in the infant. Aspirin L3 Controlled studies in breastfeeding women fail to demonstrate a risk to the infant and the possibility of harm to the breastfeeding infant is Birth Control – ONLY Acceptable remote; or the product is not orally bioavailable in an infant. L2 Safer Norethindrone, Depo-Provera, Drug which has been studied in a limited number of breastfeeding Implanon, Mirena, Plan B women without an increase in adverse effects in the infant; And/or, the evidence of a demonstrated risk which is likely to follow use of this medication in a breastfeeding woman is remote. Cetrizine (Zyrtec) L2 L3 Moderately Safe There are no controlled studies in breastfeeding women, however Dextromethorphan (Robitussin L1 the risk of untoward effects to a breastfed infant is possible; or, con- etc.) trolled studies show only minimal non-threatening adverse effects. Drugs should be given only if the potential benefit justifies the poten- Dimenhydrinate (Dramamine) L2 tial risk to the infant. L4 Possibly Hazardous Diphenhydramine (Benadryl) L2 There is positive evidence of risk to a breastfed infant or to breast- milk production, but the benefits of use in breastfeeding mothers Fluoxetine (Prozac) L2 may be acceptable despite the risk to the infant (e.g. -

Breastfeeding Is Best Booklet
SOUTH DAKOTA DEPARTMENT OF HEALTH WIC PROGRAM Benefits of Breastfeeding Getting Started Breastfeeding Solutions Collecting and Storing Breast Milk Returning to Work or School Breastfeeding Resources Academy of Breastfeeding Medicine www.bfmed.org American Academy of Pediatrics www2.aap.org/breastfeeding Parenting website through the AAP www.healthychildren.org/English/Pages/default.aspx Breastfeeding programs in other states www.cdc.gov/obesity/downloads/CDC_BFWorkplaceSupport.pdf Business Case for Breastfeeding www.womenshealth.gov/breastfeeding/breastfeeding-home-work- and-public/breastfeeding-and-going-back-work/business-case Centers for Disease Control and Prevention www.cdc.gov/breastfeeding Drugs and Lactation Database (LactMed) www.toxnet.nlm.nih.gov/newtoxnet/lactmed.htm FDA Breastpump Information www.fda.gov/MedicalDevices/ProductsandMedicalProcedures/ HomeHealthandConsumer/ConsumerProducts/BreastPumps Healthy SD Breastfeeding-Friendly Business Initiative www.healthysd.gov/breastfeeding International Lactation Consultant Association www.ilca.org/home La Leche League www.lalecheleague.org MyPlate for Pregnancy and Breastfeeding www.choosemyplate.gov/moms-pregnancy-breastfeeding South Dakota WIC Program www.sdwic.org Page 1 Breastfeeding Resources WIC Works Resource System wicworks.fns.usda.gov/breastfeeding World Health Organization www.who.int/nutrition/topics/infantfeeding United States Breastfeeding Committee - www.usbreastfeeding.org U.S. Department of Health and Human Services/ Office of Women’s Health www.womenshealth.gov/breastfeeding -

Download/Rozdzial05.Pdf (Accessed on 1 January 2007)
cancers Article Early Alcohol Use Initiation, Obesity, Not Breastfeeding, and Residence in a Rural Area as Risk Factors for Breast Cancer: A Case-Control Study Dorota Anna Dydjow-Bendek * and Paweł Zagozd˙ zon˙ Department of Hygiene and Epidemiology, Medical University of Gdansk, 80-211 Gdansk, Poland; [email protected] * Correspondence: [email protected] Simple Summary: Breast cancer became the most common cancer globally in 2021, according to the World Health Organization. The aim of the study was to evaluate risk factors for breast cancer, such as early alcohol use initiation, obesity, breastfeeding, and place of residence. The effect of alcohol consumption by girls has been assessed in only a few studies and is not fully understood. In this study, it has been found to be associated with a higher risk of breast cancer. Our study also shed light on the incidence disparity—women were more at risk in the countryside than in the city. The results of this study should be included in the preparation of breast cancer prevention programs and also aimed at women in adolescence and early adulthood because exposures during childhood and adolescence can affect a woman’s long-term risk of breast cancer. Every effort should also be made to ensure that access to knowledge is open to all, regardless of where they live, giving all women equal opportunities. Citation: Dydjow-Bendek, D.A.; Zagozd˙ zon,˙ P. Early Alcohol Use Abstract: Initiation, Obesity, Not Breastfeeding, The aim of this study was to determine the risk factors for breast cancer in the Polish and Residence in a Rural Area as Risk population. -

Clinical Update and Treatment of Lactation Insufficiency
Review Article Maternal Health CLINICAL UPDATE AND TREATMENT OF LACTATION INSUFFICIENCY ARSHIYA SULTANA* KHALEEQ UR RAHMAN** MANJULA S MS*** SUMMARY: Lactation is beneficial to mother’s health as well as provides specific nourishments, growth, and development to the baby. Hence, it is a nature’s precious gift for the infant; however, lactation insufficiency is one of the explanations mentioned most often by women throughout the world for the early discontinuation of breast- feeding and/or for the introduction of supplementary bottles. Globally, lactation insufficiency is a public health concern, as the use of breast milk substitutes increases the risk of morbidity and mortality among infants in developing countries, and these supplements are the most common cause of malnutrition. The incidence has been estimated to range from 23% to 63% during the first 4 months after delivery. The present article provides a literary search in English language of incidence, etiopathogensis, pathophysiology, clinical features, diagnosis, and current update on treatment of lactation insufficiency from different sources such as reference books, Medline, Pubmed, other Web sites, etc. Non-breast-fed infant are 14 times more likely to die due to diarrhea, 3 times more likely to die of respiratory infection, and twice as likely to die of other infections than an exclusively breast-fed child. Therefore, lactation insufficiency should be tackled in appropriate manner. Key words : Lactation insufficiency, lactation, galactagogue, breast-feeding INTRODUCTION Breast-feeding is advised becasue human milk is The synonyms of lactation insufficiency are as follows: species-specific nourishment for the baby, produces lactational inadequacy (1), breast milk insufficiency (2), optimum growth and development, and provides substantial lactation failure (3,4), mothers milk insufficiency (MMI) (2), protection from illness. -

Maternal Intake of Cow's Milk During Lactation Is Associated with Lower
nutrients Article Maternal Intake of Cow’s Milk during Lactation Is Associated with Lower Prevalence of Food Allergy in Offspring Mia Stråvik 1 , Malin Barman 1,2 , Bill Hesselmar 3, Anna Sandin 4, Agnes E. Wold 5 and Ann-Sofie Sandberg 1,* 1 Department of Biology and Biological Engineering, Food and Nutrition Science, Chalmers University of Technology, 412 96 Gothenburg, Sweden; [email protected] (M.S.); [email protected] (M.B.) 2 Institute of Environmental Medicine, Unit of Metals and Health, Karolinska Institutet, 171 77 Stockholm, Sweden 3 Department of Paediatrics, Institute of Clinical Sciences, Sahlgrenska Academy, University of Gothenburg, 405 30 Gothenburg, Sweden; [email protected] 4 Department of Clinical Science, Pediatrics, Sunderby Research Unit, Umeå University, 901 87 Umeå, Sweden; [email protected] 5 Department of Infectious Diseases, Institute of Biomedicine, Sahlgrenska Academy, University of Gothenburg, 413 90 Gothenburg, Sweden; [email protected] * Correspondence: ann-sofi[email protected] Received: 10 November 2020; Accepted: 25 November 2020; Published: 28 November 2020 Abstract: Maternal diet during pregnancy and lactation may affect the propensity of the child to develop an allergy. The aim was to assess and compare the dietary intake of pregnant and lactating women, validate it with biomarkers, and to relate these data to physician-diagnosed allergy in the offspring at 12 months of age. Maternal diet during pregnancy and lactation was assessed by repeated semi-quantitative food frequency questionnaires in a prospective Swedish birth cohort (n = 508). Fatty acid proportions were measured in maternal breast milk and erythrocytes. Allergy was diagnosed at 12 months of age by a pediatrician specialized in allergy. -

Side-Branching in the Mammary Gland: the Progesterone–Wnt Connection
Downloaded from genesdev.cshlp.org on September 27, 2021 - Published by Cold Spring Harbor Laboratory Press PERSPECTIVE Side-branching in the mammary gland: the progesterone–Wnt connection Gertraud W. Robinson,1 Lothar Hennighausen, and Peter F. Johnson2 Laboratory of Genetics and Physiology, National Institute of Diabetes and Digestive and Kidney Diseases, National Institutes of Health, Bethesda, Maryland 20892-0822 USA; 2Eukaryotic Transcriptional Regulation Section, Regulation of Cell Growth Laboratory, National Cancer Institute-Frederick Cancer Research Development Center (FCRDC), Frederick, Maryland 21702-1201 USA The mammary gland is a derivative of the ectoderm mammary ducts during puberty and pregnancy. Their whose development begins in the embryo and progresses studies demonstrate that a nuclear signal is converted after birth. The major part of development occurs in the into a secreted signal that can control the fate of adjacent adolescent and adult animal. Hormones produced by the cells in a paracrine fashion. A genetic understanding of pituitary, the ovaries, the uterus, the placenta, and the this and other signaling pathways regulating cell growth mammary gland itself control this process. Over the past in the mammary gland will improve our ability to ma- century, surgical, biological, and genetic tools have been nipulate these processes and thus design strategies for used to gain insight into physiological and pathological prevention and treatment of breast cancer. processes in the mammary gland. Originally, endocrine ablation and reconstitution experiments provided a de- scriptive framework of the role of ovarian and pituitary Tools to investigate signaling pathways hormones (Halban 1900; Nandi 1958). These experi- Several features of the mammary gland provide unique ments demonstrated a clear requirement for the ovarian opportunities for experimental manipulations to inte- steroids estrogen and progesterone for ductal growth and grate systemic, local and cell-specific signaling path- alveolar development (Topper and Freeman 1980). -

Breastfeeding Support Credentials
Who’s Who? A glance at breastfeeding support in the United States Lactation support is often needed to help mothers initiate and continue breastfeeding. There are many kinds of help available for breastfeeding mothers including peer counselors, certified breastfeeding educators and counselors, and lactation professionals such as the International Board Certified Lactation Consultant (IBCLC®). Breastfeeding support is valuable for a variety of reasons, from encouragement and emotional support to guidance and assistance with complex clinical situations. Mothers benefit from all kinds of support, and it is important to receive the right kind at the right time. The breastfeeding support categories listed below each play a vital role in providing care to mothers and babies. Breastfeeding Support Prerequisites Training Required Scope of Practice Type • 90 hours of lactation-specific education Recognized health Provide professional, • College level health science professional or evidence based, clinical courses Professional satisfactory completion lactation management; • 300-1000 clinical practice hours (International Board Certified of collegiate level educate families, health Lactation Consultant, IBCLC®) • Successful completion of a health sciences professionals and others criterion-referenced exam offered coursework. about human lactation. by an independent international board of examiners. Certified • 20-120 hours of classroom training Provide education and • Often includes a written exam guidance for families (i.e. Certified Lactation N/A Counselor, Certified or “certification” offered by the on basic breastfeeding Breastfeeding Educator, etc. ) training organization issues. Provide breastfeeding information, Peer Personal breastfeeding • 18-50 hours of classroom training encouragement, and (i.e. La Leche League, WIC experience. Peer Counselor, etc.) support to those in their community. Copyright © 2016 by USLCA. -

MATERNAL & CHILD HEALTH Technical Information Bulletin
A Review of the Medical Benefits and Contraindications to Breastfeeding in the United States Ruth A. Lawrence, M.D. Technical Information Bulletin Technical MATERNAL & CHILD HEALTH MATERNAL October 1997 Cite as Lawrence RA. 1997. A Review of the Medical Benefits and Contraindications to Breastfeeding in the United States (Maternal and Child Health Technical Information Bulletin). Arlington, VA: National Center for Education in Maternal and Child Health. A Review of the Medical Benefits and Contraindications to Breastfeeding in the United States (Maternal and Child Health Technical Information Bulletin) is not copyrighted with the exception of tables 1–6. Readers are free to duplicate and use all or part of the information contained in this publi- cation except for tables 1–6 as noted above. Please contact the publishers listed in the tables’ source lines for permission to reprint. In accordance with accepted publishing standards, the National Center for Education in Maternal and Child Health (NCEMCH) requests acknowledg- ment, in print, of any information reproduced in another publication. The mission of the National Center for Education in Maternal and Child Health is to promote and improve the health, education, and well-being of children and families by leading a nation- al effort to collect, develop, and disseminate information and educational materials on maternal and child health, and by collaborating with public agencies, voluntary and professional organi- zations, research and training programs, policy centers, and others to advance knowledge in programs, service delivery, and policy development. Established in 1982 at Georgetown University, NCEMCH is part of the Georgetown Public Policy Institute. NCEMCH is funded primarily by the U.S. -

Estrogen and Progesterone Treatment Mimicking Pregnancy for Protection from Breast Cancer
in vivo 22 : 191-202 (2008) Review Estrogen and Progesterone Treatment Mimicking Pregnancy for Protection from Breast Cancer AIRO TSUBURA, NORIHISA UEHARA, YOICHIRO MATSUOKA, KATSUHIKO YOSHIZAWA and TAKASHI YURI Department of Pathology II, Kansai Medical University, Moriguchi, Osaka 570-8506, Japan Abstract. Early age at full-term pregnancy lowers the risk of The etiology of human breast cancer is largely unknown. breast cancer in women; lactation seems to be of marginal Genetic susceptibility, hormonal effects and environmental importance and aborted pregnancy is not associated with factors appear to be major determinants. However, known reduced risk. Although early full-term pregnancy provides genetic risk factors are present in only 10% to 15% of breast protection against breast cancer, first full-term pregnancy in cancer cases (1). Many aspects of hormonal effects confer older women appears to increase the risk. The protective effect increased risk of human breast cancer. The incidence of human of pregnancy has also been observed in rats and mice; in these breast cancer is 100-fold greater for females than for males and animals, lactation has an additive effect and interrupted female reproductive history is a consistent risk factor for pregnancy provides partial but significant protection. human breast cancer (2, 3). Studies indicate that early Pregnancy at a young age ( 3 months) is highly effective, but menarche and late menopause, both of which increase the ≤ pregnancy in older animals ( 4 months) is less effective. duration of ovarian steroid exposure, positively correlate with ≥ Parity-induced protection against mammary cancer in rodents increased risk; bilateral oophorectomy at an early age is can be reproduced by short-term treatment (approximately associated with reduced risk (4). -

Substance Use in Pregnancy
MAY 2017 SUBSTANCE USE IN PREGNANCY Prepared By: Aaron Fields, MD University of Rochester Medical Center, NYS Maternal & Infant Health Center of Excellence 265 Crittenden Blvd, Rochester, NY 14642 – Tel: 585-276-7893 – Fax: 585-461-4532 [email protected] – www.mihcoe.org Introduction ..............................................................................................................................................................2 What is Addiction? .....................................................................................................................................................3 Drugs of Abuse ..........................................................................................................................................................4 Opioids ................................................................................................................................................................................ 4 Stimulants ........................................................................................................................................................................... 4 Nicotine............................................................................................................................................................................... 5 Alcohol ................................................................................................................................................................................ 6 Marijuana ........................................................................................................................................................................... -

What Makes a Mammal a Mammal?
GRADE LEVELS: 3 - 4 CORRELATION TO NEXT GENERATION SCIENCE STANDARDS: 3-LS4-3, 4-LS1-1 TEACHER’S SKILLS/PROCESSES: observation, analysis, comparison & generalization, identification, creativity OBJECTIVE: Students will be able to identify the five charac - teristics by which mammals are determined. GUIDE UNIT ONE LESSON ONE What Makes a Mammal a Mammal? BACKGROUND Classifying animals into categories and groups based on their similarities and differences is the first step in studying and understanding their WHITE-TAILED DEER origins, development and interdependence. Mammals have the following characteristics: 1. They are covered with hair or fur. 2. They are warm-blooded (mean - ing their internal body temperature is maintained at a constant level regardless of external conditions). 3. They are usually born alive and relatively well-devel - oped, having grown inside the mother’s body in a special organ called a uterus . The time spent developing in the uterus before birth is called the gestation period and varies in length from species to species (from about 13 days in the Virginia opossum to 210 days in the white-tailed deer). 4. After birth the young are fed with milk that is pro - duced by mammary glands . 5. They have larger and more complex brains than any other group of animals. Focusing on these five characteristics will enhance the students’ awareness of and interest in mammals of Illinois. It will also provide a frame of reference for exploring the similarities and differences among members of the animal kingdom and how those characteristics relate to the environment and lifestyle of individual species. 1 Wild Mammals of Illinois, Illinois Department of Natural Resources PROCEDURE AND DISCUSSION Review the student information with the class, providing students VOCABULARY with (or inviting them to provide) examples of each of the five mam- gestation period—the length of mal characteristics. -

The Milk Supply Equation
The Making of a Milk Factory Objectives . List the major factors necessary for good milk production . Describe the role of the placenta in mammary gland development during pregnancy. Explain two ways that environmental The Making of a Milk Factory contaminants may interfere with reproductive hormones © 2019 Lisa Marasco MA, IBCLC, FILCA Hormones and Receptors The Milk Supply Equation SUFFICIENT GLANDULAR TISSUE + INTACT NERVE PATHWAYS & DUCTS + ADEQUATE HORMONES & RECEPTORS + ADEQUATE LACTATION-CRITICAL NUTRIENTS + ADEQUATE & EFFECTIVE MILK REMOVAL (Good management, effective baby) = GOOD MILK SUPPLY Directors of the process Important players by stage Hormones important to milk production Puberty Pregnancy Lactation Insulin Estrogen Estrogen Cortisol Lactogenic Progesterone Progesterone Complex HPL Prolactin Chorionic Gonadotropin Maintains metabolism; Growth Hormone Growth Hormone Thyroxine - Supports & directs IGF-1 IGF-1 PTPRF pituitary Prolactin Prolactin Insulin Insulin Regulate mammary blood Cortisol Cortisol PTHrP - flow; calcium transport and Thyroxine Thyroxine homeostasis PTHrP PTHrP Oxytocin Oxytocin - milk delivery With permission: Andaluz Waterbirth Center © Lisa Marasco 2019 1 The Making of a Milk Factory Hormone receptors 101 Hormone Receptors 201 Hormones- travel thru blood or synthesized locally Dynamic: Expressed by genes ER α, β Expressed PR α, β R Up & down regulation where and PRLR L, S Can be resistant as needed OTR H Can be hypersensitive Can be influenced positively or negatively by other hormones Receptors: Integral to hormone function, located on “target tissues,” on or in cells Can be influenced by environment The Big Construction Picture PR-A Phase I ER PR-B ERα Series of stages of organo-genesis lasting until ERβ PRLrL adulthood that are irreversible (fetal through puberty) PR-B Phase II Series of changes involving growth and secretory differentiation of the lobulo-alveolar system that are reversible Estrogen Prolactin Progesterone (Preg, lactation, involution) Horseman, N.