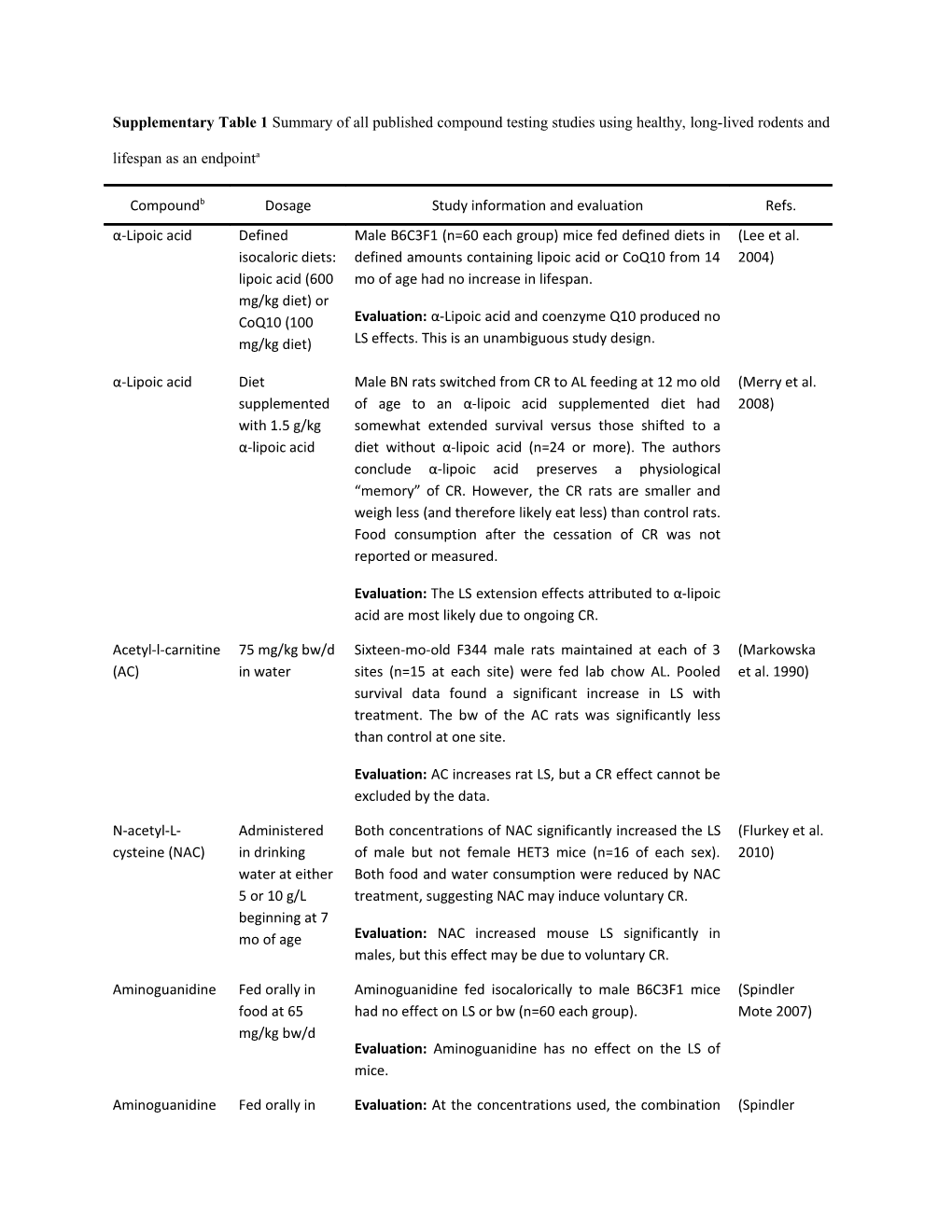
Supplementary Table 1 Summary of all published compound testing studies using healthy, long-lived rodents and lifespan as an endpointa
Compoundb Dosage Study information and evaluation Refs. α-Lipoic acid Defined Male B6C3F1 (n=60 each group) mice fed defined diets in (Lee et al. isocaloric diets: defined amounts containing lipoic acid or CoQ10 from 14 2004) lipoic acid (600 mo of age had no increase in lifespan. mg/kg diet) or CoQ10 (100 Evaluation: α-Lipoic acid and coenzyme Q10 produced no mg/kg diet) LS effects. This is an unambiguous study design.
α-Lipoic acid Diet Male BN rats switched from CR to AL feeding at 12 mo old (Merry et al. supplemented of age to an α-lipoic acid supplemented diet had 2008) with 1.5 g/kg somewhat extended survival versus those shifted to a α-lipoic acid diet without α-lipoic acid (n=24 or more). The authors conclude α-lipoic acid preserves a physiological “memory” of CR. However, the CR rats are smaller and weigh less (and therefore likely eat less) than control rats. Food consumption after the cessation of CR was not reported or measured.
Evaluation: The LS extension effects attributed to α-lipoic acid are most likely due to ongoing CR.
Acetyl-l-carnitine 75 mg/kg bw/d Sixteen-mo-old F344 male rats maintained at each of 3 (Markowska (AC) in water sites (n=15 at each site) were fed lab chow AL. Pooled et al. 1990) survival data found a significant increase in LS with treatment. The bw of the AC rats was significantly less than control at one site.
Evaluation: AC increases rat LS, but a CR effect cannot be excluded by the data.
N-acetyl-L- Administered Both concentrations of NAC significantly increased the LS (Flurkey et al. cysteine (NAC) in drinking of male but not female HET3 mice (n=16 of each sex). 2010) water at either Both food and water consumption were reduced by NAC 5 or 10 g/L treatment, suggesting NAC may induce voluntary CR. beginning at 7 mo of age Evaluation: NAC increased mouse LS significantly in males, but this effect may be due to voluntary CR.
Aminoguanidine Fed orally in Aminoguanidine fed isocalorically to male B6C3F1 mice (Spindler food at 65 had no effect on LS or bw (n=60 each group). Mote 2007) mg/kg bw/d Evaluation: Aminoguanidine has no effect on the LS of mice.
Aminoguanidine Fed orally in Evaluation: At the concentrations used, the combination (Spindler and α-lipoic acid food at 65 and of aminoguanidine and α-lipoic acid had no effect on the Mote 2007) fed together 73 mg/kg LS or bw of male B6C3F1 mice fed isocalorically with bw/d, controls (n=60 each group). respectively Evaluation: The combination of aminoguanidine and α- lipoic acid has no effect on the LS of mice.
Aminoguanidine, Aminoguanidin Evaluation: Isocaloric feeding of the combination of (Spindler α-lipoic acid, e, α- lipoic aminoguanidine, α-lipoic acid, pregnenolone and Mote 2007) pregnenolone, acid, coenzyme Q10 together had no effect on the LS or bw of and coenzyme pregnenolone, male B6C3F1 mice (n=60 each group). Q10 fed together and coenzyme Q10 at 65, 73, Evaluation: The combination of aminoguanidine, α-lipoic 0.2, and 12 acid, pregnenolone and coenzyme Q10 had no effect on mg/kg bw/d, the LS of mice. respectively, in food
Antioxidant AM fed daily AM feeding begun at 2 and 9 mo of age produced a 9.5% (Bezlepkin et mixture (AM) mixed with to 16% increase in mean and max LS of male B6 mice. Bw al. 1996) diet. See next measured monthly or bimonthly. “The results…showed column for that both experimental and control mice [were] equally details. varied in weight (data not given).” The meaning of this statement is unclear. Induced CR cannot be excluded as source of the LS increase.
AM delivered 7.5 mg β-carotene; 15 mg α-tocopherol acetate; 50 mg ascorbic acid; 25 µg sodium selenite; 38.4 mg zinc gluconate (5 mg elemental zinc); 25 mg rutin (quercetin 3 β-rutinoside)/kg bw/d
Evaluation: AM mixture increased LS significantly. However, a voluntary CR cannot be excluded as the cause.
Aspirin 21 mg/kg food Aspirin begun at 4 mo of age significantly increased the (Strong et al. (3.3 mg/kg median LS of HET3 mice by 8%. Food consumption was 2008) bw/d) not measured. The weights of the mice were described as not different than control. However, no data or analyses were shown. Study utilized large sample sizes, both sexes, and multiple testing sites.
Evaluation: Aspirin administered beginning at a younger age increased mouse LS without affecting bw. A CR effect appears unlikely.
Aspirin 21 mg/kg food Aspirin begun at 16-18 mo of age had no effect on the LS (Flurkey et al. (3.3 mg/kg of female (n=60) HET3 mice. Males were not tested. No 2010) bw/d) significant effect on bw detected (measurements made every 3 mo, and statistics reported).
Evaluation: Aspirin begun late in life has no effect on the LS of female mice.
Angiotensin- 20, 10 and 5 At all three concentrations, enalapril extended the LS of (Ferder et al. converting mg/L CF1 mice to the same extent relative to the controls 1993) enzyme (ACE) administered (n=21). This is a seriously confounded study. Survival of inhibitor in water AL. controls was only 70-75% at 10 mo, 50% at 18 mo and Enalapril 20% at 24 mo. Bw was measured only at death or at 24 mo of age. The study was discontinued at 24 mo of age, when all mice were killed. Food was administered AL, and not measured. The control mice weighed ~35 g, while all the treated groups weighed ~47 g. The mean LS reported in this study (~548 d) is much shorter than that reported elsewhere for this mouse strain (Kahn 1975).
Evaluation: The results of this study are inconclusive. Bw and its measurement are confounded. Husbandry confounds are suggested, especially in view of the studies cited below.
ACE inhibitor; Enalapril Enalapril had no effect on the median or max LS of UM- (Harrison et Enalapril administered HET3 mice. Study is robust in its use of large sample sizes, al. 2009) in food testing in both sexes, and distribution of studies over beginning at three testing sites. However, food consumption was not 120 d of age at measured. The weights of the mice were described as not 120 mg/kg different than control, but no data or analyses were food (~20 reported. mg/kg bw/d) Evaluation: At the concentration used, enalapril had no effect on LS. However, food intake was not measured and weight data and analysis are not presented.
ACE inhibitor; Enalapril Enalapril treatment produced a significant 15% decrease (Santos et al. Enalapril administered in bw gain of Wistar rats compared to control (vehicle 2009) in drinking consuming) rats (n=20). Food and water intake and bw water from 30 were determined weekly. Caloric intake was significantly d of age at 10 reduced by enalapril. At the 26 mo of age end point, the mg/kg bw/d chow and vehicle treated rats had 20% and the enalapril treated rats 45% survival.
Evaluation: At the concentration used, enalapril appeared to extend LS. A decrease in food consumption and bw suggests a CR effect is responsible.
BAPN 10 mg/d in At the concentration used, BAPN had no effect on the LS (Kohn Leash water from 40 of male or female CD*F rats. The statistical power is low (β- to 93 wk of (n=5 males and 5 females/group). Air conditioner failure 1967) aminopropiontril age, and 5 led to deaths of some mice. In some studies, higher e fumarate) mg/d concentrations of BAPN caused irreversible inhibition of thereafter growth and lowered bw.
Evaluation: At the concentration used, BAPN did not extend LS. A number of serious study confounds exist, reducing confidence in the results.
BAPN BAPN Five of six groups of LAF/J mice (n=24) treated with BAPN (LaBella administered showed an increase in mean LS by ~2 mo of treatment. Vivian 1978) at 1 or 3 Mice treated with 3 mg/mL had lower bw than controls. mg/mL in Survival curves could only be analyzed after ~40% of drinking water deaths occurred due to unexplained early deaths. beginning at 2- Voluntary CR cannot be excluded as source of LS increase. mo of age for 6, 12 or 18 mo. Evaluation: The results of a confounded study suggest BAPN may extend LS. However, voluntary CR cannot be excluded.
Boron Starting at 794 Boron in drinking water beginning at 794 d of age had no (Massie d of age, mice effect on the LS of male C57BL/6J mice. However, the 1994) were given 4.3 authors point out that the boron content of Purina #5001 ppm or 21.6 is 31.1 ppm, much higher than the amount delivered in ppm sodium the water. borate in drinking water Evaluation: A confounded study gave equivocal results.
Caffeic acid At 120 d of CAPE had no effect on the median or max LS of UM-HET3 (Harrison et phenethyl ester age, 30 and mice. Food consumption was not measured. The weights al. 2009) (CAPE) 300 mg/kg of the mice were described as not different than control, food (5 and 50 but no data or analyses were shown. Study utilized large mg/kg bw/d) sample sizes, both sexes, and multiple testing sites.
Evaluation: At the concentration used, CAPE had no effect on LS.
Centrophenoxin 0.3 g of Centrophenoxine extended the median, mean and max LS (Hochschild e centrophenoxi of Swiss Webster Albino mice (n32) by ~30%, 27% 1973) (Dimethylamino ne/L drinking (P=0.039) and 27%. However, drug treated mice were ethyl p- water from 8.6 ~11% lighter than the control mice at 22.4 mo of age, chlorophenoxyac to 18.2 mo of suggesting a possible CR effect. etatecentrophen age and from oxine 22.2 mo of age Evaluation: DMEA extends mouse LS, probably by to death inducing voluntary CR.
Coenzyme Q10 10 mg/kg Male S-D rats (n=75/group) and male C57/B17 mice (Lonnrot et (CoQ10) bw/d. CoQ was (n=43/group) were fed CoQ10 supplemented diets al. 1998) administered starting at 2 mo of age. No significant differences in LS AL in food after were found. CoQ10 levels were significantly elevated in being dissolved the plasma and liver. Animals were weighed “regularly” in soybean oil. and no differences were found. However, the weight data CoQ10 was less and their analysis were not reported. than 0.5 mg/kg/d in the Evaluation: CoQ10 supplementation has no effect on control group. mouse or rat lifespans at this dosage.
Coenzyme Q10 Defined Male B6C3F1 (n=60 each group) mice fed defined diets in (Lee et al. (CoQ10) isocaloric diet defined amounts containing lipoic acid or CoQ10 from 14 2004) with CoQ10 mo of age had no change in LS. (100 mg/kg diet) Evaluation: CoQ10 produced no LS effect at this concentration.
Coenzyme Q10 0.7 mg/kg Male Wistar rats (n=43/group) were fed AL with diets (Quiles et al. (CoQ10) bw/d CoQ10 in containing 61% polyunsaturated fatty acids. One group 2004) food was supplemented with CoQ10. CoQ10 extended mean LS by ~12% and max LS by 24%. Weights were measured and reported throughout the study, and no significant effects were found. Food intake was reported as not different than control, but no details or analyses were given. CoQ10 content was significantly higher in the liver mitochondria of supplemented rats.
Evaluation: Low dose CoQ10 supplementation of rats on a high polyunsaturated fatty acid diet appeared to extend lifespan, although a CR effect cannot be excluded.
Coenzyme Q10 Zero, 93, or Male C57BL/6 mice fed AL diets providing CoQ (Sohal et al. (CoQ10) 371 mg of supplementation beginning at 3.5 mo of age 2006) CoQ10 /kg bw (n=50/group). No significant LS difference was found fed AL in food. between groups. CoQ9 and CoQ10 in mitochondria and tissues of liver, heart, kidney, skeletal muscle, (but not brain) increased with dosage and duration of supplementation. Bw throughout study was reported, as was the method of their statistical analysis. Bw was unaffected by supplementation.
Evaluation: At the concentrations used, CoQ10 supplementation had no effect on LS or bw of male C57BL/6 mice.
Coenzyme Q10 See also in this table α-Lipoic acid and coenzyme Q10 (Lee (CoQ10) et al. 2004); and coenzyme Q10, aminoguanidine, α-lipoic acid, and pregnenolone fed together (Spindler Mote 2007).
Creatine Treated mice The mean LS of B6 mice was increased 9% and max LS (Bender et al. were fed a 3.5% by creatine (p<0.05; n=81/group). The weights of 2007) standard the mice or their food consumption were not reported or rodent diet discussed. containing 1% Creatine and Evaluation: Creatine extends the LS of mice, but induced, control mice voluntary CR cannot be excluded. were fed a standard diet.
Cysteine 0.5 and 1% in AKRc male (n=30) and C3Hd (n=29) female mice were fed (Harman hydrochloride food AL two supplemented diets from 1.5 to 16 mo of age. For 1957) AKR mice, 1% cysteine produced a 20% increase in the age at which 50% of the mice were dead (from ~8 to ~10 mo; P≤0.01). No effect was found with C3H mice. AKR and C3H mice are short-lived strains. Weights were measured monthly and reported. No analysis of the weights was presented. Controls consumed two different amounts of food.
Evaluation: Cysteine may have extend the LS of AKR mice. However, a CR effect on LS cannot be excluded. The study is confounded by differential caloric intake among the control mice and unanalyzed weight data.
Cysteine 1% in food AKR male (n=42), C3H female (n=49) and Swiss male (Harman hydrochloride (n=48) mice were fed supplemented diets AL from 1961) weaning to 16 mo of age. The half-survival time of AKR mice was prolonged 14.5% (P < 0.1). However, cysteine had a slight adverse effect on the LS of C3H and Swiss mice, although it is unclear whether this effect was statistically significant. AKR and C3H mice are short-lived strains. Average weight was determined monthly, and reported. No analysis or discussion of the weights was presented.
Evaluation: Cysteine hydrochloride may have extend the LS of the short-lived AKR mouse strain, but shortened that of C3H and Swiss mice. These results are consistent with those from of previous study. A CR effect on LS cannot be excluded.
Cysteine 0.5 and 1% in Male LAFl mice were fed AL a custom, semi-synthetic diet (Harman hydrochloride food with or without the drug from of 6 to 8 wks of age 1968) (control n=120; treated n=60). The drug produced no change in mean LS. The study is confounded by an increase in mortality induced by the semi-synthetic diet which the decreased 50% survival of controls from 20 to 15 mo of age relative to chow diet; and an increase in mortality at about 14 mo of age due to “poor animal care”.
Evaluation: In a multiply confounded study, cysteine hydrochloride had no effect on the LS of a long-lived mouse strain.
Dehydro- 25 μg/mL No increase in LS was detected in DHEAS treated mice fed (Pugh et al. epiandrosterone DHEAS in their control or CR diets. Food consumption and weights were 1999) sulfate (DHEAS) drinking water monitored. DHEAS did not affect bw or cancer patterns. AL Evaluation: At the concentration used, DHEAS has no effect on LS.
Deltaran Deltaran Max (17.1%; P<0.01; n=54) but not mean LS increased for (Popovich et [mixture of injected at 100 female SHRe mice subcutaneously injected with Deltaran. al. 2003) synthetic δ-sleep mg/kg bw for 5 The mean LS of the treated mice was not increased. No inducing peptide consecutive data on food consumption or bw given. (DSIP; Trp-Ala- d/mo Gly-Gly-Asp-Ala- Evaluation: Deltaran extends max LS, but an induced, Ser-Gly-Glu) and voluntary CR effect cannot be excluded. glycine (w/w, 1:10)]
Deprenyl [a.k.a. Oral (1.0 mg Oral deprenyl significantly increased LS in female Syrian (Stoll et al. (-)Deprenyl; selegiline/kg hamsters (~20% increase; p=0.02; n=35), but not in 1997; Stoll et Selegiline; food); equaling males. Female control hamsters had a shorter LS than al. 1994) Carbex; about 0.05 mg male controls, and deprenyl equalized their LS. Selegiline Eldepryl)] selegiline/kg can be used orally in Syrian hamsters because they do not bw/d degrade 90% of it in the first pass through the liver as do mice and humans. No effect of selegiline on bw was found. Food intake and bw were measured regularly, and these data were shown.
Evaluation: Oral deprenyl extends the LS of female Syrian hamsters.
Deprenyl [a.k.a. 0.25 mg/kg s.c. Deprenyl increased the LS of castrated female Wistar rats (Dallo 2001) (-)Deprenyl; three times a by almost double (P<0.05; n=9), but not intact rats. No Selegiline; wk. Age at food consumption or bw data. Carbex; which the Eldepryl)] treatment Evaluation: Castration together with deprenyl increased started is not the LS of female rats. Induced, voluntary CR cannot be given. excluded.
Deprenyl [a.k.a. Dissolved in Deprenyl extended mean and max LS in male F344 rats (Kitani et al. (-)Deprenyl; saline and (n=12/group). Rats received deprenyl starting at 18 mo of 2006; Kitani Selegiline; given by s.c. age for 13 mo. Mean LS significantly increased by 8.4% et al. 2005) Carbex; injection. at (0.25 mg/kg) and 5.6% (0.5 mg/kg). Rats given 1.0 mg/kg Eldepryl)] 0.25, 0.5, and die sooner than controls. Rats were fed AL. 1.0 mg/kg/injectio Evaluation: Injected deprenyl extended the LS of male n, three times rats. However, an induced CR effect cannot be excluded. a wk. No information regarding food consumption or bw was provided.
Deprenyl [a.k.a. Mice received Deprenyl and Dinh lang root extract extended the mean (Yen Knoll (-)Deprenyl; DLRE (10 LS of OFA-l mice (by 35 and 43%, respectively) and 1992) Selegiline; mg/kg bw together their effects were greater (64%). Max LS was Carbex; oral), deprenyl similarly extended. The drugs did not suppress food Eldepryl)] and (0.25 mg/kg bw intake or change bw. Dinh lang s.c.), DLRE plus (Polycias deprenyl, or a Evaluation: Oral deprenyl and Dinh lang root extract fruticosum L.) saline control extends the LS of male OFA-l mice. root extract three times/wk (DLRE) alone and beginning at together 12-mo-old
2,2'- 0.5 and 1% in AKR male (n=25 or 30) and C3H (n=29) female mice were (Harman Diaminodiethyl food fed AL the two diets from 2 to 16 mo of age. For AKR 1957) disulfide mice, a 20% increase in “half-survival time” was found dihydrochloride with 0.5% DDS (from ~8 to ~10 mo; P≤0.01). No effect (DDS) was found with C3H mice. Weights were measured monthly and reported. No analysis of the weights was presented. Controls consumed two different amounts of food.
Evaluation: DDS may have extend the LS of AKR mice. However a CR effect on LS cannot be excluded. The study is confounded by possible differential caloric intake among the control mice and unanalyzed weight data.
2,2'- 1% in food AKR male (n=43), C3H (n=48) female and Swiss male (Harman Diaminodiethyl (n=51) mice were fed the supplemented diet AL from 1961) disulfide after weaning to 16 mo of age. 1% DDS somewhat dihydrochloride shortened the LS of all 3 strains of mice, including that of (DDS) AKR, which had experienced an increase in LS when fed DDS at 0.5% in food (see above) (Harman 1957). Average weight was determined monthly, and was reported. No analysis or discussion of the weights was presented.
Evaluation: 1% DDS in food shortens the lifespan of mice.
Digoxin (Trade 12.5 mg Digoxin increased mean LS 29% in male and 14% in (Coburn et names: Lanoxin; Digoxin / kg of female mice. The survival curves were rectangular. The al. 1974) Digitek; and food. control vs. treated males had average bw of 31.1 and 28.2 Lanoxicaps) g; the control and treated females weights of 25.9 and 21.5 g. The hearts weighed less in the treated groups.
Evaluation: Digoxin treatment extends mouse LS, apparently by inducing voluntary CR.
Dinitrophenol Administered DNP treatment increased median LS 6.7% and mean LS by (Caldeira da (DNP) orally in 7.1% (p=0.038). Food consumption was measured and did Silva et al. drinking water not differ from control. Bw was reduced ~10%. 2008) (~1 mg/L DNP) Evaluation: DNP slightly extended median and mean LS without changing food consumption. However, bw was decreased, perhaps due to its well-described effect of uncoupling oxidative phosphorylation.
2,6-Di-tert-butyl 0.25 and 0.5% Male LAFl mice were fed AL a custom, semi-synthetic diet (Harman hydroquinone in food from of 6 to 8 wks of age (control n=120; treated n=60). 1968) The drug produced no change in mean LS. The study is confounded by an increase in mortality induced by the semi-synthetic diet, which the decreased 50% survival of controls from 20 to 15 mo of age relative to a chow diet; and an increase in mortality at about 14 mo of age due to “poor animal care”.
Evaluation: In a multiply confounded study, 2,6-di-tert- butyl hydroquinone had no effect on the LS of mice.
2,6-Di-tert-butyl- 0.25 and 0.5% Male LAFl mice were fed AL a custom, semi-synthetic diet (Harman 4-methylphenol in food from of 6 to 8 wks of age (control n=120; treated n=60). 1968) (BHT; butylated Both concentrations of BHT increased the mean LS (by hydroxytoluene) 45% for 0.50% BHT; P<0.01). But, BHT produced a 5-12% reduction in bw, suggesting a CR effect on LS. The studies were confounded by an increase in mortality induced by the custom, semi-synthetic diet, which the decreased 50% survival of controls from 20 to 15 mo of age relative to a chow diet; and an increase in mortality at about 14 mo of age due to “poor animal care”.
Evaluation: In a multiply confounded study, BHT extended the mean LS of mice. However, it reduced bw dose responsively, suggesting a CR effect on LS. The synthetic diet used induced early mortality, and poor animal care led to increased mortality at 14 months of age.
2,6-Di-tert-butyl- BHT at 0.75% Male BALB/c mice fed AL, BHT containing food from 8 or (Clapp et al. 4-methylphenol in Purina 11 weeks of age had statistically significant increases in 1979) (BHT; butylated Laboratory mean survival relative to controls. The BHT consuming hydroxytoluene) mice were “generally heavier”, although no data are Chow shown, and no information regarding weighing of the mice is given. Livers of treated mice had “increased liver size” at necropsy. No data were shown. The increase in weight is in contrast to the other BHT studies reviewed herein.
Evaluation: BHT may have increased the mean LS of BALB/c mice. A CR effect appears unlikely, although relevant data are not shown. Treatment beginning at 8 vs 11 weeks of age was less effective at extending LS.
2,6-Di-tert-butyl- MEA 1% in Weanling and retired breeder female B6 mice were fed (Kohn 1971) 4-methylphenol diet. BHT 0.2% MEA and BHT supplemented diets AL. Neither produced (BHT; butylated to 0.5% in clear difference in mean or max LS. Animals that received hydroxytoluene) food. MEA and BHT did not attain control weights. and MEA (2- mercaptoethyla Evaluation: MEA and BHT had no effect on mean or max mine) LS.
DMEA (a.k.a. Administered DMEA extended mean remaining life by 50% (P=0.018) (Hochschild Deanol; orally in water; and max LS by 11% when administered to ~21 mo old 1973) dimethylethanol 86 μg of male A/J mice (n=57 each group). However, the treated amine; N,N- dimethylamino group had 5% lower bw than controls after treatment. dimethyl-2- ethanol Aminoethanol; acetamidobenz Evaluation: DMEA extends mean and max LS. Induced, β- oate/mL voluntary CR is likely responsible. dimethylaminoet (equivalent to hyl alcohol; β- ~7 mg hydroxyethyldim dimethylamino ethylamine). A ethanol/kg hydrolysis bw/d product of centrophenoxine
DOPA [L-dopa Fed 1.0 or 40 Four to 5 wk-old, male Swiss albino mice fed L-DOPA (Cotzias et al. (levodopa, L-3,4- mg Ldopa/kg weighed less and lived longer. The LS study was 1974) dihydroxyphenyl bw in food terminated at 19 mo of age. The survivors were no Ldopa, alanine)] (ground Purina 39%; 1 mg/g bw, 38%; 40 mg/g bw, 73% (P < 0.001). After chow) AL 3 wk, “differences in food consumption among surviving animals tended to disappear; body weight tended to show a gain”.
Evaluation: Oral DOPA extends LS and at least initially reduces bw and food intake. Whether DOPA extends LS in the absence of CR is difficult to judge from the data given.
DOPA [L-dopa L-dopa in 5 mg Male Swiss Albino mice (n=100/group) treated with 1.0, (Cotzias et al. (levodopa, L-3,4- increments 20, and 40 mg/g diet L-dopa starting from 4-5 wk of age dihydroxyphenyl from 1 to 100 had a dose dependant increase in median and mean LS. 1974) alanine)] mg/g diet fed Food consumption was reported as not significantly AL. The max different. However, a significant decrease in weight was concentration reported in treated mice. During 1st mo of treatment, the that caused no 40 mg/g diet group weighed 66.2 ± 7.6% (mean ± SD) of deaths was 40 control bw. mg/g diet for males and 80 Evaluation: DOPA extends LS, reduces bw, but does not mg/g diet for change food consumption. DOPA may induce a females. discordance between bw and food consumption. See earlier study above (Cotzias et al. 1974).
Echinacea The extract Echinacea purpurea root extract in food may have (Brousseau purpurea root was fed in the increased the LS of BALB/cByJ male mice eating AL. The Miller 2005) extract chow at 2 LS curves of treated and untreated crossed at about 300 d mg/mouse/d of age. The authors arbitrarily chose a point when the from 7 wk until curves diverged to end the experiment. At this time more just beyond 13 Echinacea-consuming mice were alive. The authors do mo of age not report accurate measurements of food consumption or bw.
Evaluation: Echinacea purpurea may increase the survival of mice, although a CR effect and a technical bias in the results cannot be excluded.
Emoxipine [2- 100-150 mg/kg Two mo old female SHK mice (n=112, control=118), C3HA (Emanuel ethyl-6-methyl- (method of mice (n=90, control=170), AKR mice (n=50, control=50), Obukhova
3- delivery and and (C57BL X CBA)F1 mice (n=200, control=200) were 1978) hydroxypyridine diet used were treated with 100-150 mg/kg emoxipine in an unspecified hydrochloride] not specified). manner. Mean and max LS of treated C3HA and SHK mice
were increased significantly. in AKR and (C57BLxCBA)F1 mice there was no effect on LS. No bw or food intake data were provided. The method of drug delivery and the diet used are not specified. The article references Russian language journals which are not readily available.
Evaluation: Emoxipine extends the LS of C3HA, and SHK mice. However induced, voluntary CR cannot be excluded. This study would be difficult to repeat.
Enterosorption - Administered Male Wistar rats fed a sorbent containing diet starting at (Frolkis et al. dietary sorbent in 10 d courses 28 mo of age had a significant increase in mean and max 1989; Frolkis (SKN sorbent: at 1 mo LS. Food intake and bw were not reported. These data et al. 1984) non-coated intervals in a appear to be published in two different places (Frolkis et nitrogen- dosage of 10 al. 1984; Frolkis et al. 1989). containing ml/kg bw Evaluation: Entrosorption extended the LS of male Wistar carbon) rats, but induced, voluntary CR cannot be excluded as the cause.
Epithalamin The dosages Epithalamin injection on 5 consecutive d/mo increased (Anisimov et (low-molecular are difficult to the mean LS of outbred female rats (strain not specified) al. 1992) weight pineal interpret as is by 6.2%. A citation describing the preparation and peptide the composition of Epithalamin, in Russian was not readily preparation) composition of available. No information regarding food intake or bw Epithalamin was presented. (see Study information Evaluation: Epithalamin may have increased rat LS, and however induced voluntary CR cannot be excluded as the evaluation) cause. This study would be difficult to repeat.
Epithalone (a Subcutaneous Group 1 female LIO rats were maintained on a 12h (Vinogradova synthetic Ala- injections of dark:light cycle (LD); Group 2, were maintained on et al. 2007) Glu-Asp-Gly 0.1 μg illumination mimicking that at Petrozavodsk State peptide) Epithalone, 5 University (NL); and Group 3 were maintained on times a wk in constant light (LL) (n=40 to 55/group). Half of each of the 0.1 ml saline 3 groups received Epithalon or vehicle. Epithalone had no beginning at 4 effect on mean LS. Epithalone reduced the max LS of LD mo of age. rats but increased the max LS of NL and LL rats relative to LD rats. No information regarding food intake or bw was given.
Evaluation: Epithalone may increase the max LS of photoperiod stressed rats, although a CR effect cannot be excluded because neither food intake nor bw were reported.
Epithalone (a Subcutaneous Male LIO rats were maintained on 3 different dark:light (Vinogradova synthetic Ala- injections of regimen (see Study information and evaluation of et al. 2008) Glu-Asp-Gly 0.1 μg (Vinogradova et al. 2007), above. Half of each group peptide) Epithalone, 5 received Epithalon. The peptide had no significant effect times a wk in on mean or max LS. No information regarding food intake 0.1 ml saline or bw were presented. beginning at 4 mo of age. Evaluation: Epithalone does not increase the LS of control or photoperiod stressed male rats. These results appear at odds with those cited immediately above.
Exercise and Exercise AL and Sulfamerazine (a sulfa antibiotic) extended the LS of rats (Sperling et sulfamerazine fed (strain not specified) significantly (~20%). The treated rats al. 1978) Sulfamerazine were 11% (males) and 36% (females) heavier than at 250 mg/kg in controls. Sulfamerazine treated, exercising rats were diet considerably heavier than controls. Food consumption was not measured. Evaluation: Sulfamerazine extended rat LS. However, its effects, and the effects of exercise on bw are counterintuitive. Sulfamerazine may reduce energy expenditure or increase food consumption and extend lifespan. Alternatively, the drug may have mitigated a bacterial pathogen in the animal colony. n-6/n-3 Fatty Control chow No effect on LS was found when 10 mo old (n=12-14), (Valencak acid ratio diet and chow female C57BL/6 mice were fed diets containing either n-6 Ruf 2010) diet enriched enriched or n-3 enriched chow. The control group with sunflower weighed significantly less than the treated mice. oil (n-6 rich; 750 g/6 kg Evaluation: Differences in the ratio of n-6/n3 fatty acids pellets) or do not appear to influence mouse LS. salmon oil (n-3 rich; 750 g/6 kg pellets) was fed AL. n-6/n-3 Fatty Food was Three groups of 33 Wistar rats were fed the various food (Takeuchi et acid ratio adjusted to an containing fatty acids with different n-6 to n-3 ratios AL al. 2009) n-6/n-3 ratio of from 4 wk of age. There were no significant differences either 1, 4, or mean life span, food intake, or bw among the groups. 16, but with a constant Evaluation: Variations in n-6/n-3 Fatty acid ratios do not polyunsaturate alter mouse LS. d to monounsaturat ed to saturated fatty acid ratio.
Flumazenil 3 to 4 mg/kg/d Flumazenil treatment of F344 rats from 13 to 25 mo of (Marczynski (a.k.a. in drinking age (n=20/group) significantly increased the age to which et al. 1994) flumazepil, Ro water acidified 16 of the rats in each group survived by approximately 2 15-1788, to pH 3.0) mo. No food consumption or bw data were reported. Anexate, Lanexat, Evaluation: Flumazenil may have increased the LS of rats, Mazicon, but induced, voluntary CR cannot be excluded as the Romazicon) cause.
Fluoro-deprenyl 6-mo old male Injection of fluoro-deprenyl (n = 40) or saline (n = 20) (Dallo Knoll [(-)p-fluoro- Wistar rats three times a wk for 25 mo resulted in 3 of 20 saline- 1992) deprenyl] treated with treated and 15 of 40 drug-treated males surviving to 24 0.01 mg/kg bw, mo of age (p=0.05). No food consumption or weight data s.c. were reported. (p-Fluoro-L-deprenyl is a halogenated derivative of L-deprenyl which is almost indistinguishable in potency and pharmacology) Evaluation: Fluoro-deprenyl may have extended the LS of Wistar rats, but induced, voluntary CR cannot be excluded as the means.
Ginkgo biloba Beginning at 2 GE significantly increased male F344 rat max LS by 8%. (Winter extract (GE) (EGb mo of age GE Caloric intake was controlled to maintain adult bw at 1998) 761; “most was ~300 g. There was no significant difference in weight widely used form administered between the treated and untreated control groups at the of Ginkgo biloba orally mixed time of death. in clinical with 3 mL of studies… sweetened Evaluation: GE extend the max LS of rats. An induced standardized in condensed voluntary CR effect cannot be excluded by the data given, its content of milk vehicle at but appears unlikely given the feeding procedures used. ginkgo flavone a dose of 50 glycosides and mg/kg bw?f terpenoids”
Green tea Administered The average LS of polyphenol treated male B6 mice was (Kitani et al. polyphenols in water (80 significantly longer than that of control mice (6.4% 2007) mg/L) increase; n=50 mice/group). Max LS was not significantly beginning at 13 different. No significant differences in bw among groups mo of age. were found.
Evaluation: Green tea polyphenols may increase the mean LS of mice. An induced voluntary CR effect cannot be excluded but appears unlikely.
High fat or high High fat or high A high fat diet decreased average longevity. Results were (French et al. carbohydrate carbohydrate highly significant in male and less significant in female 1953) diet diets fed AL. rats (strain not identified). Two experimental diets, one high in carbohydrate and one high in fat were fed AL. Food consumption was measured and reported (n=15/group). High fat fed rats consumed 4.8 and 5.7% less total calories (males and females, respectively).
Evaluation: A high fat diet reduces rat LS.
Hydroxylamine 1% and 2% in AKR male (n=42), C3H female (n=49), and Swiss male (Harman hydrochloride food (n=48) mice were fed the supplemented diets AL from 1961) weaning to 16 mo of age. The “half survival time” of AKR mice was increased 17% by 2% hydroxylamine (P<0.05). The other strains were unaffected. AKR and C3H mice are short-lived strains while Swiss is a long-lived strain. Average weights were determined monthly, and reported. No analysis or discussion of the weights was presented.
Evaluation: Hydroxylamine extend the LS of the short- lived AKR mouse strain. However, hydroxylamine had no effect on the LS of the normal, long lived Swiss mouse. A CR effect on LS cannot be excluded by the data shown.
Hydroxylamine 1 and 2% in Male LAFl mice were fed AL a semi-synthetic diet (Harman hydrochloride food formulated with or without the drug from of 6 to 8 wks of 1961) age (control n=120; treated n=60). The drug produced no change in mean LS. The study is confounded by an increase in mortality induced by the semi-synthetic diet, which the decreased 50% survival of controls from 20 to 15 mo of age; and an increase in mortality at about 14 mo of age due to “poor animal care”.
Evaluation: In a multiply confounded study, hydroxylamine hydrochloride had no effect on the LS of mice.
Isoflavone soy Low and high Mouse LS was increased by a low isoflavone diet in two (Bartke et al. protein diets soy protein strains of normal mice. A high isoflavone diet increased LS 2004) (Low vs. high) diets were 32 in one strain of normal mice but reduced the LS of substituted for and 972 mg/kg another strain. Diets were fed AL. Weights were different casein diets diet fed AL in the groups fed the different diets.
Evaluation: Soy isoflavones may extend the LS of some strains of mice, but the effects of induced voluntary CR cannot be excluded.
Kombucha Treatment oral Kombucha consumption from weaning until death (Hartmann et 15% solution of significantly increased the LS of B6 mice by ~5% (males; al. 2000) kombucha n=15) and 2% (females; n=17). Mice were fed AL. beverage AL; Kombucha significantly decreased the weight of both control sexes. Signs of organ damage were found in treated mice. received water. Evaluation: Kombucha increased mouse LS, but induced voluntary CR appears the probable reason.
Marine collagen MCPs MCPs were administered to groups of 40, 5 wk old S-D (Liang et al. peptides (MCPs; administered (equal numbers of males and females). Bw and food 2010) a protease in food at 2.25, consumption were measured weekly to bi-weekly. Mean hydrolysate of 4.5, or 9% LS was significantly increased by MCPs. Bw and their chum salmon (wt/wt), with statistical analysis were given, and there were no skin collagen) proportionally significant differences. There were no significant resuced diet differences in food consumption between the groups. protein. Evaluation: MCPs extend the mean LS of S-D rats.
MEA (2- MEA 1% in Weanling and retired breeder female B6 mice were fed (Kohn 1971) mercaptoethyla diet. BHT 0.2% MEA and BHT supplemented diets AL. Neither produced a mine) and BHT to 0.5% in diet. clear difference in mean or max LS. Animals that received (butylated MEA and BHT did not attain control weights. hydroxytoluene) Evaluation: MEA and BHT have no effect on mean or max LS.
Melatonin Fed orally in Melatonin (n=60 each group) had no effect on the LS or (Spindler food at 41 bw of male B6C3F1 mice fed isocalorically with controls. Mote 2007) µg/kg bw/d Evaluation: Melatonin did not effect the longevity of male B6C3F1 mice.
Mercaptoethan 0.5% in food AKR male (n=25) and C3H female (n=28) mice consuming (Harman ol (2- a drug containing diet AL from 2.5 to 16 mo of age 1957) mercaptoethan experienced no increase in “half-survival time”. Weights ol; 2-ME) were measured monthly and reported. No analysis of the weights was presented.
Evaluation: 2-ME had no effect on the LS of AKR or C3H mice. The study is confounded by possible differential caloric intake among the control mice and unanalyzed weight data.
Mercaptoethano Administered 2-ME extended the mean and max LS of male BC3F1 mice (Heidrick et l (2- orally in food (n=60/group) by ~12% and ~13%, respectively (p<0.005). al. 1984) mercaptoethano AL at 0.25% of Reported bw of the two groups were statistically different l; 2-ME) diet (w/w) at only one time point. Food consumption was not measured.
Evaluation: 2-ME extended the mean and max LS of mice while producing little effect on bw. A CR effect cannot be excluded, but appears unlikely.
2- 0.5 and 1% in AKR male (n=22 or 28) and C3H female (n=25 or 28) mice (Harman Mercaptoethyla food were fed AL the two diets from 1 or 2 to 16 mo of age. 1957) mine For AKR mice, a 20% increase in “half-survival time” was hydrochloride found with 1% 2-mercaptoethylamine (from ~8 to ~10 (MEA) mo; P≤0.01). No effect was found with C3H mice. Weights were measured monthly and reported. No analysis of the weights was presented.
Evaluation: Cysteine may have extend the LS of AKR but not C3H mice. However, a CR effect on LS cannot be excluded. The study is confounded by possible differential caloric intake among the control mice and unanalyzed weight data.
2- 1% in food AKR male (n=42), C3H female (n=49) and Swiss male (Harman Mercaptoethyla (n=48) mice were fed the supplemented diet AL from 1961) mine after weaning to 16 mo of age. The “half-survival time” of hydrochloride C3H mice was extended from 14.5 to 18.3 months (26%; (MEA) P<0.01), in contrast to the previous study (Harman 1957). Also in contrast to the previous study, MEA had no effect on the half-survival time of AKR mice. MEA had no effect on the LS of Swiss mice, a long-lived mouse strain. AKR and C3H mice are short-lived strains. Average weight was determined monthly, and reported. No analysis or discussion of the weights was presented.
Evaluation: MEA may have extend the LS of the short- lived C3H mouse strain. However, these results contrast with a previous study by the same author. MEA had no effect on the LS of the normal, long lived Swiss mouse strain. A CR effect on LS cannot be excluded.
2- 0.5 & 1% in Male LAFl mice were fed AL a commercial chow or a (Harman Mercaptoethyla food custom semi-synthetic diet from of 6 to 8 wks of age 1968) mine (control n=120; treated n=60). No statistically significant hydrochloride change in LS was found using the commercial chow. With (MEA) the synthetic diet, 1% MEA produced a 12% increase in mean LS (P<0.01). But, 0.5 and 1% MEA produced a 6 and 12% reduction in bw in these mice, suggesting a CR effect on LS. The studies also were confounded by an increase in mortality induced by the semi-synthetic diet, which the decreased 50% survival of controls from 20 to 15 mo; and an increase in mortality at about 14 mo of age due to “poor animal care”.
Evaluation: In a multiply confounded study, MEA extended the mean LS of mice. However, it reduced bw dose responsively, suggesting a CR effect on LS, the synthetic diet induced early mortality, and poor animal care led to increased mortality at 14 months of age.
Metformin 100 mg/kg in Metformin extended the mean LS of female SHR mice by (Anisimov et drinking water ~38% and max LS by ~21%. Mice were fed AL. Metformin al. 2008b) “slightly modified the food consumption” and “decreased (estimated bw after the age of 20 mo”. dosage of ~10 mg/kg bw/d) Evaluation: Metformin extended mean and max LS, but an induced, voluntary CR effect appears possible, even likely.
Metformin 300 mg/kg F344 rats were fed a diet containing metformin or a (Smith, Jr. et bw/d in their control diet AL. A control group was pair fed with the al. 2010) food metformin treated group. Bw and food intake were measured weekly. Metformin did not significantly increase mean or max LS.
Evaluation: Metformin had no effect on the LS of F344 rats.
N- Both given in C3H mice (n=20/group) treated with the combination of (Oxenkrug et acetylserotonin drinking water NAS and melatonin had a ~20% increase in LS (males) (p< al. 2001) (NAS) combined at 2.5 mg/kg 0.01). No affect was found for females. Some of the with melatonin bw/d. studies were right censored. Weights were not reported Melatonin and for the LS mice. However, in other studies of B6 mice NAS stock reported herein, NAS and melatonin reduced the weights solutions 10 of the mice, suggesting food intake may have been mg/mL in 1% reduced. Tween-20. Controls given Evaluation: NAS and melatonin in combination increased solvent. the lifespan of male C3H mice, but a CR effect appears probable.
Nitroflurbiprofen 200 mg/kg NFP begun at 4 mo of age had no effect on the median or (Strong et al. (NFP) food (33 mg/kg max LS of HET3 mice. Study utilized large sample sizes, 2008) bw/d) both sexes, and multiple testing sites. However, food consumption was not measured. The weights of the mice were described as not different than control. However, it is unclear whether these observations are anecdotal or systematic.
Evaluation: NFP at this concentration has no effect on mouse LS.
Nitroflurbiprofen 200 mg/kg NFP begun at 16-18 mo of age had no effect on the LS of (Flurkey et al. (NFP) food (33 mg/kg female HET3 mice (n=60; males not tested). No significant 2010) bw/d) effect on bw was detected (measurements made every 3 mo, and statistics reported).
Evaluation: NFP at this concentration has no effect on the LS of female mice.
Nordihydroguaia 2.5 g of NDGA begun at 9 mo of age significantly increased the (Strong et al. retic acid (NDGA) NDGA/kg of median but not max LS of male (but not female) UM- 2008) food (~417 HET3 mice by 12%. No effect found on max LS. Food mg/kg bw/d) consumption was not measured. The weights of the mice were described as not different than control. Study utilized large sample sizes, both sexes, and multiple testing sites.
Evaluation: NDGA increased mouse LS significantly without changing mouse weights. A potential CR effect cannot be excluded. This dose of NDGA significantly decreased the weight of isocalorically fed male B6C3F1 mice (Fig. 1).
Nordihydroguaia 2.5 g of NDGA begun at 16-18 mo of age had no effect on the LS (Flurkey et al. retic acid (NDGA) NDGA/kg of of female UM-HET3 mice (n=60; males were not tested). 2010) food (~417 Food was given AL. No significant effect on bw was mg/kg bw/d) detected (measurements made every 3 mo, and statistics reported).
Evaluation: NDGA begun at 17-18 mo of age had no effect on the LS of female mice.
N-tert-butyl-α- 0.25 mg/mL of PBN treatment begun at 24.5 mo of age (n=50 each (Saito et al. phenylnitrone PBN in drinking group) prolonged mean and max LS of male B6 mice by 1998) (PBN) water about 3% (p<0.005). PBN administration beginning at 18.5 or 21.5 mo produced no LS extension. There was no effect on bw, which was measured throughout study and reported. Food consumption was not measured.
Evaluation: PBN appears to modestly extend mean and max LS of male mice. An induced voluntary CR effect cannot be excluded, but appears unlikely.
N-tert-butyl-α- 350 mg/kg 4-OH-PBN begun at 4 mo of age had no effect on the (Strong et al. phenylnitrone food (~53 median or max LS of HET3 mice. Study utilized large 2008) derivative 4-OH- mg/kg bw/d) sample sizes, both sexes, and multiple testing sites. The α-phenyl-N-tert- weights of the mice were described as not different than butyl nitrone (4- control, but no data or analyses were shown. OH-PBN) Evaluation: 4-OH-PBN at this dose had no effect on LS.
N-tert-butyl-α- 315 mg/kg 4-OH-PBN begun at 16-18 mo of age had no effect on the (Flurkey et al. phenylnitrone food (~52 LS of female UM-HET3 mice (n=60/group; males were not 2010) derivative 4-OH- mg/kg bw/d) tested). No significant effect on bw was detected, α-phenyl-N-tert- measurements made every 3 mo, and statistics were butyl nitrone (4- reported. OH-PBN) Evaluation: 4-OH-PBN at this dose has no effect on the LS of female mice.
Nucleic acids An unspecified Five control (untreated) and 5 rats of an unspecified (Odens 1973) volume of a strain began treatment at 750 d of age. The diet and its chloroform manner of administration was not described. All the saturated untreated rats died before 900 d. Four of the treated rats water solution died between the ages of 1600 and 1900 d, and one lived containing DNA 2250 d. No statistical analyses were reported. The at 3 treated rats were reported to have gained 1.5-2.3 gm, but mg/mL)and weights were not reported. “ordinary” RNA at an Evaluation: Too little information is given and too many unspecified confounds exist in this report for the study to be concentration repeated or evaluated productively. administered weekly by injection in an unspecified manner
Phenformin 2 mg/d Chronic treatment of female C3H/Sn mice prolonged (Dilman administered mean LS by 23% and decreased spontaneous tumor Anisimov in 0.2 ml of tap incidence. Food intake and bw were not measured. 1980) water/mouse/ d os 5 Evaluation: Phenformin extended the LS of female mice times/wk of this relatively short lived strain, although an induced beginning at CR effect cannot be excluded. It is possible that these 3.5 mo of age. studies were performed at the time that C3H mice were infected with mouse mammary tumor virus.
Pineal factor Subcutaneous PF administered to groups (n=30-41) of female Swiss- (Anisimov et extract (PF) injections 5 derived outbred SHR mice beginning at 3.5 mo (young) or al. 1989) d/mo of 0.1 mg 12 mo (middle age) extended average and max LS “of PF or normal tumor-free young mice” by 20% and by 17% for middle saline from 3.5 aged. No information regarding food intake or bw was or 12 mo of reported. The composition and method of preparation of age. PE are published in difficult to obtain Russian language journals. The studies would be difficult to repeat.
Evaluation: PF extends mouse LS, although an induced voluntary CR effect cannot be excluded. This study would be difficult to repeat.
Piperoxane Beginning at 16 Piperoxane extended mean survival of F344 rats by (Compton et mo. 16.7%. This is a weakly powered (n=8) preliminary study. al. 1995) intraperitoneal The methods section refers to a paper for which no injections 3 record could be found. Food was given AL. Food mg/kg bw/d consumption was not investigated. The article states piperoxane or there was no significant difference in bw, but no data are saline every 48 given. hours for four mo. Evaluation: Piperoxane may extend rat LS, but an induced, voluntary CR effect cannot be excluded.
Pregnenolone Fed orally in At the concentrations used pregnenolone and melatonin (Spindler and melatonin food at 200 together (n=60 each group) had no effect on the LS or bw Mote 2007) µg/kg bw/d of male B6C3F1 mice fed isocalorically. and 41 µg/kg bw/d, Evaluation: The concentrations of pregnenolone and respectively melatonin used had no effect on the longevity of mice. Propyl gallate 0.25 & 0.5% in Male LAFl mice were fed AL a semi-synthetic diet (Harman food formulated with or without the drug from of 6 to 8 wks of 1968) age (control n=120; treated n=60). The drug produced no change in mean LS. The study is confounded by an increase in mortality induced by the semi-synthetic diet, which the decreased 50% survival of controls from 20 to 15 mo of age; and an increase in mortality at about 14 mo of age due to “poor animal care”.
Evaluation: In a multiply confounded study, propyl gallate had no effect on the LS of mice.
Rapamycin Microencapsul Rapamycin significantly extended the LS of HET3 mice by (Harrison et ated rapamycin an average of 12.7% for females and 9.3% for males. Max al. 2009) administered LS also was increased significantly. However, food at 600 d of age consumption was not measured. The weights of the mice at 14 mg/kg were described as not different than control, but no data food (~2.24 mg or analysis were shown. A confound in the pooled male of data is that groups which were treated with rapamycin at rapamycin/kg 600 d already had lower mortality than the groups which body wt/d). served as controls. In a later publication, many of these authors report that this concentration of rapamycin reduced the bw of HET3 mice (Miller et al. 2010) (see below). Study utilized large sample sizes, both sexes, and multiple testing sites.
Evaluation: Microencapsulated rapamycin increased mouse LS significantly. Weight data and analysis were not shown. A later report by many of these same investigators suggests that rapamycin decreases the weight of this strain of mouse. Thus, induced, voluntary CR cannot be excluded, and may be a likely source of the longevity effects.
Rapamycin Rapamycin was Rapamycin administered from the age of 9 mo extended (Miller et al. administered the max and median LS of AL fed HET3 mice by 10% for 2010) in food (14 males and 18% for females. The treatment reduced the mg/kg) weight of the mice by ~10% in males and 6% in females.
Evaluation: Rapamycin may extend the LS of UM-HET3 mice, however a CR effect on LS cannot be excluded, and appears likely.
Reduced NIH-31 open C57BL6 male mice were isocalorically fed Regular or Low (Cai et al. advanced formula diets diets from 4 mo of age. Food consumption and bw were 2007) glycation end were prepared monitored regularly and reported. Mice fed the Low diet product (AGE) identically had a 15% and 6% extension of median and max LS. The containing diet except one was exposed to 70 Low diet had a 50% reduction in AGEs. to 75°C for 1-2 m and 55°C for Evaluation: LS was extended by the Low AGE diet. 30 m, as per normal (Regular); the other was prepared by exposure to 80°C for 1 m (Low).
Resveratrol Resveratrol in Resveratrol fed at 3 concentrations beginning at 12 mo of (Pearson et food, age had no effect on the LS of male B6 mice consuming a al. 2008) approximate standard diet AL or EOD. doses in mg/kg bw/d were: for Evaluation: Resveratrol at low to high doses has no effect standard diet, on the LS of mice fed a normal diet or fed EOD. 7.9, 30.9 and 204; for EOD diet, 7.6 and 30.4
Resveratrol 300 and 1200 Resveratrol administered from the age of 9 mo had no (Miller et al. ppm food (50 effect on the LS of AL fed, male and female HET3 mice. 2010) and 200 mg/kg No data regarding weight or food consumption given. bw/d) Evaluation: Resveratrol at low to high doses has no effect on the LS of mice.
Simvastatin 12 and 120 Simvastatin administered from the age of 9 mo had no (Miller et al. mg/kg food; 2 effect on the LS of AL fed, male and female HET3 mice. 2010) and 20 mg/kg No data regarding weight or food consumption were bw/d given. What was reported as not different than control.
Evaluation: Simvastatin at a low and a high dose has no effect on the LS of mice fed a normal diet.
SkQ1 SkQ1 in Administration of SkQ1 from 2 mo of age to female, (Anisimov et (Mitochondria- drinking water outbred SHR mice may have extend mean but not max LS al. 2008a) targeted fresh each d at at 5 nmol/kg bw/d. No evidence for statistical analysis of plastoquinone a dosage of these data could be found. Food was given AL, but the derivative) 0.5, 5, or 50 authors state that food consumption and bw were not nmol SkQ1/kg significantly different between groups. No methods, data, bw/d or their analysis were given. SHR mice are a normal, but short lived strain. Evaluation: SkQ1 may have extended mean LS of SHR mice, but a lack of statistical analysis and methods information undermine confidence in the results. The possibility of an induced, voluntary CR effect cannot be excluded.
Sodium chloride From weaning Elevated NaCl had no effect on the LS of Albino rats (Lee 1963) in diet 0.51 mEq Na (n=30) fed a basic diet AL. (0.03% NaCl); 9.96mEq Evaluation: Enhanced NaCl concentrations had no effect Na(0.58% on rat LS. NaCl); and 24.60 mEq Na(1.45% NaCl)/100 g of diet.
Tetrahydrocurcu 0.2% in diet B6 mice (n=50/group) which started a diet containing (Kitani et al. min (w/w) tetrahydrocurcumin at 13 mo of age had ~12% (P<0.01) 2007) longer average LS than control mice. Max LS was 6.5% greater (P<0.01). There was a decrease in bw of 2 to 4% (P < 0.05) in the treated mice.
Evaluation: Tetrahydrocurcumin increased LS, but an induced, voluntary CR effect appears likely.
Thioproline (L- 2.0 g/kg of Thioproline induced a 20% decrease in spontaneous food (Navarro et thiazolidine-4- food from 28 intake, a 10% decrease in bw at 100 wk of age, and a 23- al. 2007) carboxylic acid) wk of age 29% increase in median and max LS.
Evaluation: LS extension by thioproline was likely induced by voluntary CR.
Thymic peptides Dosages and TM4 treated CFW mice had a 25% increase in LS. CFW, (Ghanta et and fragments of routes of A/HEN and B6 mice were treated with thymic tissue al. 1990) thymic administration extract, FTS, TP5, TM4 and TH beginning at 2 mo of age. hormones, not specified. This article is a two pages in length, and no details are thymic humoral Article states given. The composition or purification methods for the factor (FTS), they were extracts are not provided. Neither food intake nor bw thymopentin given weekly were reported. (TP5), TM4 (“an starting from 2 enzyme resistant mo of age. Evaluation: TM4 increased the survival of CFW mice, but variant of FTS”) the effects of induced, voluntary CR cannot be excluded. and thymosin These studies could not be repeated with the information (TH) and thymic given in the publication. tissue extracts. Thymogen Thymogen: Five-mo-old outbred female LIO rats injected with (Anisimov et (synthetic L-Glu- subcutaneously thymogen (5 μg/rats, n=44; control, n=32) 5 times/wk for al. 2000) L-Trp) injected 12 mo had an increase in max LS (P<0.001), but no dissolved in 0.2 change in mean LS. The publication states there was no ml of saline 5 change in bw, but no data are shown, and food times/wk for consumption was not measured. 12 mo Evaluation: Thymogen extended the max LS of female LIO rats, possibly without altering bw. However, a potential voluntary CR effect cannot be excluded.
Thymus Factor Subcutaneous A peptide isolated from thymus, TF, administered to (Anisimov et (TF) injections 5 female Swiss-derived outbred SHR mice (n=30-41) al. 1989) d/mo of 5 μg beginning at 3.5 mo (young) or 12 mo (middle age) TF in normal increased LS “of tumor-free young mice” by 12% (P < saline from 3.5 0.05) and the LS of middle-aged animals by 15% (P <0.05). or 12 mo of No information regarding caloric intake or bw is given. age. The composition and method of preparation of TF is published in difficult to obtain Russian language journals. The studies would be difficult to repeat.
Evaluation: TF extended mouse LS of female SHR mice, although induced, voluntary CR cannot be excluded as the mechanism. This study would be difficult to repeat.
Vigconic 28 (VI- Fed AL in diet Treatment of male and female C57BL/6J mice (n=41–60) (Ko et al. 28; A Chinese at 0.05 and significantly extended median LS by ~10%. Effects on bw 2010) herbal formula) 0.5% (wt/wt; are described as “moderate”. Food intake is not reported. 50-80 mg/kg (VI-28 is composed of Radix Ginseng, Cornu Cervi, bw) starting at Cordyceps, Radix Salviae, Semen Allii, Fructus Cnidii, 36 wk of age Fructus Evodiae, and Rhizoma Kaempferiae.) until death Evaluation: VI-28 extended the LS male and female C57BL/6J mice. But, voluntary, induced CR can not be excluded, and appears likely.
Vilon (synthetic Vilon: 0.1 μg Vilon extend the max LS of female CBA mice (n=60). (Khavinson L-Lys-L-Glu) Vilon Treated survivors at 22 and 23 mo were ~1.2- and 2.6- et al. 2000; subcutaneously fold more numerous than controls (p<0.01). The mean LS Khavinson injected/mous was not increased. No information was given regarding Anisimov e beginning at food intake or weight. 2000) 6-mo of age for 5 d/mo. Evaluation: Vilon increased max LS of female CBA mice, Control but an induced, voluntary CR effect cannot be excluded. injected with saline.
Vitamin C 2% in food AKR male (n=27) and C3H (n=28) female mice were fed an (Harman (ascorbic acid) ascorbic acid containing diet AL from 1 or 2.5 to 16 mo of 1957) age. No increase in the age at which 50% of the mice are dead was found. Weights were measured monthly and reported. No analysis of the weights was presented.
Evaluation: Vitamin C did not extend the LS of AKR or C3H mice. The study is confounded by possible differential caloric intake among the control mice and unanalyzed weight data.
Vitamin C 1% ascorbic Vitamin C at the concentration used increased the (Massie et al. (ascorbic acid) acid (1,430 average LS of B6 male mice by ~9% (P<0.05). Max LS 1984) mg/kg bw) in increased 2.9%. The ascorbic acid treated mice weighed 6 drinking water to 7% less than the control mice. Food intake was not controlled or reported.
Evaluation: Vitamin C increased the LS of mice, but a CR effect appears likely.
Vitamin E (RRR- 2 or 200 mg of Vitamin E, at the concentration used, had little or no (Porta et al. α-tocopherol) dl-α- significant effect on the LS of male Wistar rats. Vitamin E 1980) tocopherol fed was the only antioxidant present in the diets. to rats consuming a Evaluation: Vitamin E at the concentration used had little diet with 15% if any effect on the LS of mice regardless of the degree of coconut oil saturation of the fats in their diets. (saturated fat diet), safflower oil (unsaturated fat diet) or a combination of both (CFD)
Vitamin E (RRR- dl-α- A slight increase in the LS of vitamin E supplemented LAF1 (Blackett Hall α-tocopherol) tocopherol, 25 [(C57L x A)F1 hybrid] mice was reported. 1981) mg/kg diet, Supplementation shortened the LS of C3H mice. Food sprayed over intake was not measured. The study reported there was the food as a no difference in the bw of treated mice. However, no solution in data or analysis are given. Studies were discontinued at diethyl ether. 28 mo of age.
Evaluation: Vitamin E may have strain-specifically extended or shortened mouse lifespan. Induced, voluntary CR cannot be excluded as the reason for the increase in LS.
Vitamin E (RRR- 20, 40 and 400 Vitamin E, at the concentration used, had no effect on the (Morley α-tocopherol) mg/kg of LS of female Balb/c mice (n=42 and 45/group). Mice were Trainor 2001) vitamin E in fed vitamin E diets (apparently) from weaning. Neither standard chow, weights nor food consumption were reported. The age at fed AL. which treatment was begun was not reported.
Standard chow Evaluation: Vitamin E had no effect on the LS of female contains 40 Balb/c mice at the concentration tested. mg/kg vitamin E.
Vitamin E (RRR- Vitamin E (500 Vitamin E, glutathione, melatonin, and strawberry extract (Meydani et α-tocopherol), ppm), combined had no effect on LS at the concentrations al. 1998) glutathione, glutathione tested. Male C57BL\6 mice were fed AL supplemented melatonin, and (0.5%); vitamin AIN-76 diet beginning at 18 mo of age. Mice were killed strawberry E plus at 24 mo of age or when 50% of the mice had died. No glutathione effect on mean LS was found. The authors state, “all the extract (500 ppm + mice in this study were heavier than expected, no 0.5%, difference in weight was observed between dietary respectively),m groups”, which seems contradictory. elatonin (11 ppm), and Evaluation: Vitamin E, glutathione, melatonin, and strawberry strawberry extract had no effect on the mean LS of male extract (1%). C57BL\6 mice at the concentration tested. Orally in diet. aThe author reviewed only English language publications for this review. bAbbreviations: AL, Ad libitum; B6, C57BL/6; bw, body weight; d, day or days; EOD, every other day;
F344, Fischer 344; LS, lifespan; max, maximum; min, minutes; mo, month or months; os, by mouth; wk, week or weeks; S-D, Sprague-Dawley. cAKR mice are a normal strain with a short LS (~50% survival at 300 d). dC3H mice are a short lived strain with a high breast cancer incidence. During the era in which many of these studies were performed they were infected with a vertically transmitted mammary tumor virus. eSHR mice are a normal strain with a short LS (~50% survival at 350 d). fThe notation “mg/kg bw?” indicates that the article did not make clear whether it was reporting dose/kg diet or dose/kg body weight. References
Anisimov VN, Bakeeva LE, Egormin PA, Filenko OF, Isakova EF, Manskikh VN, Mikhelson VM, Panteleeva AA,
Pasyukova EG, Pilipenko DI, Piskunova TS, Popovich IG, Roshchina NV, Rybina OY, Saprunova VB,
Samoylova TA, Semenchenko AV, Skulachev MV, Spivak IM, Tsybul'ko EA, Tyndyk ML, Vyssokikh MY,
Yurova MN, Zabezhinsky MA, Skulachev VP (2008a) Mitochondria-targeted plastoquinone derivatives as
tools to interrupt execution of the aging program. 5. SkQ1 prolongs lifespan and prevents development of
traits of senescence. Biochemistry (Mosc ) 73:1329-1342
Anisimov VN, Berstein LM, Egormin PA, Piskunova TS, Popovich IG, Zabezhinski MA, Tyndyk ML, Yurova MV,
Kovalenko IG, Poroshina TE, Semenchenko AV (2008b) Metformin slows down aging and extends life span
of female SHR mice. Cell Cycle 7:2769-2773
Anisimov VN, Bondarenko LA, Khavinson VK (1992) Effect of pineal peptide preparation (epithalamin) on life span
and pineal and serum melatonin level in old rats. Ann N Y Acad Sci 673:53-57
Anisimov VN, Khavinson VK, Morozov VG (2000) Immunomodulatory synthetic dipeptide L-Glu-L-Trp slows down
aging and inhibits spontaneous carcinogenesis in rats. Biogerontology 1:55-59
Anisimov VN, Loktionov AS, Khavinson VK, Morozov VG (1989) Effect of low-molecular-weight factors of thymus
and pineal gland on life span and spontaneous tumour development in female mice of different age.
Mech Ageing Dev 49:245-257
Bartke A, Peluso MR, Moretz N, Wright C, Bonkowski M, Winters TA, Shanahan MF, Kopchick JJ, Banz WJ (2004)
Effects of Soy-derived diets on plasma and liver lipids, glucose tolerance, and longevity in normal, long-
lived and short-lived mice. Horm Metab Res 36:550-558
Bender A, Beckers J, Schneider I, Holter SM, Haack T, Ruthsatz T, Vogt-Weisenhorn DM, Becker L, Genius J, Rujescu
D, Irmler M, Mijalski T, Mader M, Quintanilla-Martinez L, Fuchs H, Gailus-Durner V, de Angelis MH, Wurst W, Schmidt J, Klopstock T (2007) Creatine improves health and survival of mice. Neurobiol Aging 29:1404-
1411
Bezlepkin VG, Sirota NP, Gaziev AI (1996) The prolongation of survival in mice by dietary antioxidants depends on
their age by the start of feeding this diet. Mech Ageing Dev 92:227-234
Blackett AD, Hall DA (1981) Vitamin E--its significance in mouse ageing. Age Ageing 10:191-195
Brousseau M, Miller SC (2005) Enhancement of natural killer cells and increased survival of aging mice fed daily
Echinacea root extract from youth. Biogerontology 6:157-163
Cai W, He JC, Zhu L, Chen X, Wallenstein S, Striker GE, Vlassara H (2007) Reduced oxidant stress and extended
lifespan in mice exposed to a low glycotoxin diet: Association with increased AGER1 expression. Am J
Pathol 170:1893-1902
Caldeira da Silva CC, Cerqueira FM, Barbosa LF, Medeiros MH, Kowaltowski AJ (2008) Mild Mitochondrial
Uncoupling in Mice Affects Energy Metabolism, Redox Balance and Longevity. Aging Cell 7:552-560
Clapp NK, Satterfield LC, Bowles ND (1979) Effects of the antioxidant butylated hydroxytoluene (BHT) on mortality
in BALB/c mice. J Gerontol 34:497-501
Coburn AF, Grey RM, Rivera SM (1974) Observations on the relation of heart rate, life span, weight and
mineralization in the digoxin-treated A-J mouse. Johns Hopkins Med J 128:169-193
Compton DM, Dietrich KL, Smith JS (1995) Influence of the alpha 2 noradrenergic antagonist piperoxane on
longevity in the Fischer-344 rat: A preliminary report. Psychol Rep 77:139-142
Cotzias GC, Miller ST, Nicholson AR, Jr., Maston WH, Tang LC (1974) Prolongation of the life-span in mice adapted
to large amounts of L-dopa. Proc Natl Acad Sci U S A 71:2466-2469
Dallo J (2001) Effect of longevity treatment with (-)deprenyl on lifespan and sexual behavior in female rats.
Neurobiology (Bp) 9:33-35 Dallo J, Knoll J (1992) Effect of (-)-para-fluoro-deprenyl on survival and copulation in male rats. Acta Physiol Hung
79:125-129
Dilman VM, Anisimov VN (1980) Effect of treatment with phenformin, diphenylhydantoin or L-dopa on life span
and tumour incidence in C3H/Sn mice. Gerontology 26:241-246
Emanuel NM, Obukhova LK (1978) Types of experimental delay in aging patterns. Exp Gerontol 13:25-29
Ferder L, Inserra F, Romano L, Ercole L, Pszenny V (1993) Effects of angiotensin-converting enzyme inhibition on
mitochondrial number in the aging mouse. Am J Physiol 265:C15-C18
Flurkey K, Astle CM, Harrison DE (2010) Life Extension by Diet Restriction and N-Acetyl-L-Cysteine in Genetically
Heterogeneous Mice. J Gerontol A Biol Sci Med Sci 65A:1275-1284
French CE, Ingram RH, Uram JA, Barron GP, Swift RW (1953) The influence of dietary fat and carbohydrate on
growth and longevity in rats. J Nutr 51:329-339
Frolkis VV, Nikolaev VG, Bogatskaya LN, Stupina AS, Shcherbitskaya EV, Kovtun AI, Paramonova GI, Sabko VE,
Shaposhnikov VM, Rushkevich Y (1984) Enterosorption in prolonging old animal lifespan. Exp Gerontol
19:217-225
Frolkis VV, Nikolaev VG, Paramonova GI, Shchorbitskaya EV, Bogatskaya LN, Stupina AS, Kovtun AI, Sabko VE,
Shaposhnikov VM, Muradian KK (1989) Effect of enterosorption on animal lifespan. Biomater Artif Cells
Artif Organs 17:341-351
Ghanta VK, Hiramoto NS, Hiramoto RN (1990) Thymic peptides as anti-aging drugs: Effect of thymic hormones on
immunity and life span. Int J Neurosci 51:371-372
Harman D (1957) Prolongation of the normal life span by radiation protection chemicals. J Gerontol 12:257-263
Harman D (1961) Prolongation of the normal lifespan and inhibition of spontaneous cancer by antioxidants. J
Gerontol 16:247-254 Harman D (1968) Free radical theory of aging: effect of free radical reaction inhibitors on the mortality rate of male
LAF mice. J Gerontol 23:476-482
Harrison DE, Strong R, Sharp ZD, Nelson JF, Astle CM, Flurkey K, Nadon NL, Wilkinson JE, Frenkel K, Carter CS, Pahor
M, Javors MA, Fernandez E, Miller RA (2009) Rapamycin fed late in life extends lifespan in genetically
heterogeneous mice. Nature 460:392-395
Hartmann AM, Burleson LE, Holmes AK, Geist CR (2000) Effects of chronic kombucha ingestion on open-field
behaviors, longevity, appetitive behaviors, and organs in C57BL/6 mice: A pilot study. Nutrition 16:755-
761
Heidrick ML, Hendricks LC, Cook DE (1984) Effect of dietary 2-mercaptoethanol on the life span, immune system,
tumor incidence and lipid peroxidation damage in spleen lymphocytes of aging BC3F1 mice. Mech Ageing
Dev 27:341-358
Hochschild R (1973) Effect of dimethylaminoethyl p-chlorophenoxyacetate on the life span of male Swiss Webster
Albino mice. Exp Gerontol 8:177-183
Kahn AJ (1975) Changes in ethanol consumption by C3H and CF1 mice with age. J Stud Alcohol 36:1107-1123
Khavinson VK, Anisimov VN (2000) A synthetic dipeptide vilon (L-Lys-L-Glu) inhibits growth of spontaneous tumors
and increases life span of mice. Dokl Biol Sci 372:261-263
Khavinson VK, Anisimov VN, Zavarzina NY, Zabezhinskii MA, Zimina OA, Popovich IG, Shtylik AV, Malinin VV,
Morozov VG (2000) Effect of vilon on biological age and lifespan in mice. Bull Exp Biol Med 130:687-690
Kitani K, Kanai S, Miyasaka K, Carrillo MC, Ivy GO (2005) Dose-dependency of life span prolongation of F344/DuCrj
rats injected with (-)deprenyl. Biogerontology 6:297-302 Kitani K, Kanai S, Miyasaka K, Carrillo MC, Ivy GO (2006) The necessity of having a proper dose of (-)deprenyl (D) to
prolong the life spans of rats explains discrepancies among different studies in the past. Ann N Y Acad Sci
1067:375-382
Kitani K, Osawa T, Yokozawa T (2007) The effects of tetrahydrocurcumin and green tea polyphenol on the survival
of male C57BL/6 mice. Biogerontology 8:567-573
Ko KM, Chiu PY, Leung HY, Siu AH, Chen N, Leong EP, Poon MK (2010) Long-term dietary supplementation with a
yang-invigorating Chinese herbal formula increases lifespan and mitigates age-associated declines in
mitochondrial antioxidant status and functional ability of various tissues in male and female C57BL/6J
mice. Rejuvenation Res 13:168-171
Kohn RR (1971) Effect of antioxidants on life-span of C57BL mice. J Gerontol 26:378-380
Kohn RR, Leash AM (1967) Long-term lathyrogen administration to rats, with special reference to aging. Exp Mol
Pathol 7:354-361
LaBella F, Vivian S (1978) Beta-aminopropiontrile promotes longevity in mice. Exp Gerontol 13:251-254
Lee CK, Pugh TD, Klopp RG, Edwards J, Allison DB, Weindruch R, Prolla TA (2004) The impact of alpha-lipoic acid,
coenzyme Q10 and caloric restriction on life span and gene expression patterns in mice. Free Radic Biol
Med 36:1043-1057
Lee KY (1963) Some effects of high and low sodium intake on a vegetarian diet in rats. I. Longevity, food
consumption, weight gain, organ weight, serum and bone sodium and potassium, sodium and potassium
balance and hematology. Yonsei Med J 4:6-16
Liang J, Pei XR, Wang N, Zhang ZF, Wang JB, Li Y (2010) Marine collagen peptides prepared from chum salmon
(Oncorhynchus keta) skin extend the life span and inhibit spontaneous tumor incidence in Sprague-
Dawley Rats. J Med Food 13:757-770 Lonnrot K, Holm P, Lagerstedt A, Huhtala H, Alho H (1998) The effects of lifelong ubiquinone Q10 supplementation
on the Q9 and Q10 tissue concentrations and life span of male rats and mice. Biochem Mol Biol Int
44:727-737
Marczynski TJ, Artwohl J, Marczynska B (1994) Chronic administration of flumazenil increases life span and protects
rats from age-related loss of cognitive functions: A benzodiazepine/GABAergic hypothesis of brain aging.
Neurobiol Aging 15:69-84
Markowska AL, Ingram DK, Barnes CA, Spangler EL, Lemken VJ, Kametani H, Yee W, Olton DS (1990) Acetyl-1-
carnitine. 1: Effects on mortality, pathology and sensory-motor performance in aging rats. Neurobiol
Aging 11:491-498
Massie HR (1994) Effect of dietary boron on the aging process. Environ Health Perspect 102 Suppl 7:45-48
Massie HR, Aiello VR, Doherty TJ (1984) Dietary vitamin C improves the survival of mice. Gerontology 30:371-375
Merry BJ, Kirk AJ, Goyns MH (2008) Dietary lipoic acid supplementation can mimic or block the effect of dietary
restriction on life span. Mech Ageing Dev 129:341-348
Meydani M, Lipman RD, Han SN, Wu D, Beharka A, Martin KR, Bronson R, Cao G, Smith D, Meydani SN (1998) The
effect of long-term dietary supplementation with antioxidants. Ann N Y Acad Sci 854:352-360
Miller RA, Harrison DE, Astle CM, Baur JA, Boyd AR, de Cabo R, Fernandez E, Flurkey K, Javors MA, Nelson JF,
Orihuela CJ, Pletcher S, Sharp ZD, Sinclair D, Starnes JW, Wilkinson JE, Nadon NL, Strong R (2010)
Rapamycin, But Not Resveratrol or Simvastatin, Extends Life Span of Genetically Heterogeneous Mice. J
Gerontol A Biol Sci Med Sci 66:191-201
Morley AA, Trainor KJ (2001) Lack of an effect of vitamin E on lifespan of mice. Biogerontology 2:109-112 Navarro A, Sanchez-Pino MJ, Gomez C, Bandez MJ, Cadenas E, Boveris A (2007) Dietary thioproline decreases
spontaneous food intake and increases survival and neurological function in mice. Antioxid Redox Signal
9:131-141
Odens M (1973) Prolongation of the life span in rats. J Am Geriatr Soc 21:450-451
Oxenkrug G, Requintina P, Bachurin S (2001) Antioxidant and antiaging activity of N-acetylserotonin and melatonin
in the in vivo models. Ann N Y Acad Sci 939:190-199
Pearson KJ, Baur JA, Lewis KN, Peshkin L, Price NL, Labinskyy N, Swindell WR, Kamara D, Minor RK, Perez E,
Jamieson HA, Zhang Y, Dunn SR, Sharma K, Pleshko N, Woollett LA, Csiszar A, Ikeno Y, Le Couteur D, Elliott
PJ, Becker KG, Navas P, Ingram DK, Wolf NS, Ungvari Z, Sinclair DA, de Cabo R (2008) Resveratrol Delays
Age-Related Deterioration and Mimics Transcriptional Aspects of Dietary Restriction without Extending
Life Span. Cell Metab 8:157-168
Popovich IG, Voitenkov BO, Anisimov VN, Ivanov VT, Mikhaleva II, Zabezhinski MA, Alimova IN, Baturin DA,
Zavarzina NY, Rosenfeld SV, Semenchenko AV, Yashin AI (2003) Effect of delta-sleep inducing peptide-
containing preparation Deltaran on biomarkers of aging, life span and spontaneous tumor incidence in
female SHR mice. Mech Ageing Dev 124:721-731
Porta EA, Joun NS, Nitta RT (1980) Effects of the type of dietary fat at two levels of vitamin E in Wistar male rats
during development and aging. I. Life span, serum biochemical parameters and pathological changes.
Mech Ageing Dev 13:1-39
Pugh TD, Oberley TD, Weindruch R (1999) Dietary intervention at middle age: Caloric restriction but not
dehydroepiandrosterone sulfate increases lifespan and lifetime cancer incidence in mice. Cancer Res
59:1642-1648
Quiles JL, Ochoa JJ, Huertas JR, Mataix J (2004) Coenzyme Q supplementation protects from age-related DNA
double-strand breaks and increases lifespan in rats fed on a PUFA-rich diet. Exp Gerontol 39:189-194 Saito K, Yoshioka H, Cutler RG (1998) A spin trap, N-tert-butyl-alpha-phenylnitrone extends the life span of mice.
Bioscience, Biotechnology & Biochemistry 62:792-794
Santos EL, de Picoli SK, da Silva ED, Batista EC, Martins PJ, D'Almeida V, Pesquero JB (2009) Long term treatment
with ACE inhibitor enalapril decreases body weight gain and increases life span in rats. Biochem
Pharmacol 78:951-958
Smith DL, Jr., Elam CF, Jr., Mattison JA, Lane MA, Roth GS, Ingram DK, Allison DB (2010) Metformin
supplementation and life span in Fischer-344 rats. J Gerontol A Biol Sci Med Sci 65:468-474
Sohal RS, Kamzalov S, Sumien N, Ferguson M, Rebrin I, Heinrich KR, Forster MJ (2006) Effect of coenzyme Q10
intake on endogenous coenzyme Q content, mitochondrial electron transport chain, antioxidative
defenses, and life span of mice. Free Radical Biology and Medicine 40:480-487
Sperling GA, Loosli JK, Lupien P, McCay CM (1978) Effect of sulfamerazine and exercise on life span of rats and
hamsters. Gerontology 24:220-224
Spindler SR, Mote PL (2007) Screening Candidate Longevity Therapeutics Using Gene-Expression Arrays.
Gerontology 53:306-321
Stoll S, Hafner U, Kranzlin B, Muller WE (1997) Chronic treatment of Syrian hamsters with low-dose selegiline
increases life span in females but not males. Neurobiol Aging 18:205-211
Stoll S, Hafner U, Pohl O, Muller WE (1994) Age-related memory decline and longevity under treatment with
selegiline. Life Sci 55:2155-2163
Strong R, Miller RA, Astle CM, Floyd RA, Flurkey K, Hensley KL, Javors MA, Leeuwenburgh C, Nelson JF, Ongini E,
Nadon NL, Warner HR, Harrison DE (2008) Nordihydroguaiaretic acid and aspirin increase lifespan of
genetically heterogeneous male mice. Aging Cell 7:641-650 Takeuchi H, Sekine S, Noguchi O, Murano Y, Aoyama T, Matsuo T (2009) Effect of life-long dietary n-6/n-3 fatty acid
ratio on life span, serum lipids and serum glucose in Wistar rats. J Nutr Sci Vitaminol (Tokyo) 55:394-399
Valencak TG, Ruf T (2010) Feeding into old age: Long-term effects of dietary fatty acid supplementation on tissue
composition and life span in mice. J Comp Physiol B 181:289-298
Vinogradova IA, Bukalev AV, Zabezhinski MA, Semenchenko AV, Khavinson VK, Anisimov VN (2007) Effect of Ala-
Glu-Asp-Gly peptide on life span and development of spontaneous tumors in female rats exposed to
different illumination regimes. Bull Exp Biol Med 144:825-830
Vinogradova IA, Bukalev AV, Zabezhinski MA, Semenchenko AV, Khavinson VK, Anisimov VN (2008) Geroprotective
effect of ala-glu-asp-gly peptide in male rats exposed to different illumination regimens. Bull Exp Biol Med
145:472-477
Winter JC (1998) The effects of an extract of Ginkgo biloba, EGb 761, on cognitive behavior and longevity in the rat.
Physiol Behav 63:425-433
Yen TT, Knoll J (1992) Extension of lifespan in mice treated with Dinh lang (Policias fruticosum L.) and (-)deprenyl.
Acta Physiol Hung 79:119-124