Floristic Analysis of the Southwestern United States
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Latin America and Caribbean Region LIST of ACRONYMS
Inclusive and Sustainable Industrial Development in Latin America and Caribbean Region LIST OF ACRONYMS ALBA Bolivarian Alliance for the Americas IPs Industrial Parks BIDC Barbados Investment and Development INTI National Institute of Industrial Corperation Technologies (Argentina) BRICS Brazil, Russian Federation, India, China ISID Inclusive and Sustainable Industrial and South Africa („emerging economies“) Development CAF Development Bank for Latin America ITPOs Investment and Technology Promotion CAIME High Level Centre for Research, Offices Training and Certification of Production LATU Technological Laboratory of Uruguay (Uruguayan Project) MERCOSUR Southern Common Market CAN Andean Community MoU Memorandum of Understanding CARICOM Caribbean Community ODS Ozone Depleting Substances CELAC Community of Latin American and OESC Organization of Eastern Caribbean States Caribbean States OFID OPEC Fund for International Development CFCs Chloro-Fluoro-Carbons PCBs Poly-Chlorinated Biphenyls CIU Uruguayan Chamber of Industries POPs Persistent Organic Pollutants CNI National Confederation of Brazil PPPs Public Private Partnerships COPEI Peruvian Committee on Small Industry RO Regional Office ECLAC Economic Commission for Latin America SDGs Sustainable Development Goals EU European Union SELA Latin American Economic System FAO Food and Agriculture Organization (UN SEZs Special Economic Zones System) SICA Central American Integration System GEF Global Environmental Facility SMEs Small and Medium-sized Enterprises GNIC Great Nicaraguan Interoceanic -
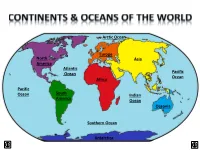
North America Other Continents
Arctic Ocean Europe North Asia America Atlantic Ocean Pacific Ocean Africa Pacific Ocean South Indian America Ocean Oceania Southern Ocean Antarctica LAND & WATER • The surface of the Earth is covered by approximately 71% water and 29% land. • It contains 7 continents and 5 oceans. Land Water EARTH’S HEMISPHERES • The planet Earth can be divided into four different sections or hemispheres. The Equator is an imaginary horizontal line (latitude) that divides the earth into the Northern and Southern hemispheres, while the Prime Meridian is the imaginary vertical line (longitude) that divides the earth into the Eastern and Western hemispheres. • North America, Earth’s 3rd largest continent, includes 23 countries. It contains Bermuda, Canada, Mexico, the United States of America, all Caribbean and Central America countries, as well as Greenland, which is the world’s largest island. North West East LOCATION South • The continent of North America is located in both the Northern and Western hemispheres. It is surrounded by the Arctic Ocean in the north, by the Atlantic Ocean in the east, and by the Pacific Ocean in the west. • It measures 24,256,000 sq. km and takes up a little more than 16% of the land on Earth. North America 16% Other Continents 84% • North America has an approximate population of almost 529 million people, which is about 8% of the World’s total population. 92% 8% North America Other Continents • The Atlantic Ocean is the second largest of Earth’s Oceans. It covers about 15% of the Earth’s total surface area and approximately 21% of its water surface area. -

State of the Region: Asia Pacific
kefk State of the region: Asia Pacific March 2021 Economy GDP growth, selected countries Business confidence - manufacturing PMIs % change on a yr ago 2020 Q2 2020 Q3 2020 Q4 2020 50=no change, seasonally adjusted 60 India -7.0 -24.4 -7.3 0.4 Japan -4.9 -10.3 -5.8 -1.3 55 Indonesia -2.1 -5.3 -3.5 -2.2 50 Korea -0.9 -2.8 -1.1 -1.2 45 Australia -2.4 -6.3 -3.7 -1.1 40 Thailand -6.2 -12.0 -6.4 -4.2 Malaysia -5.6 -17.1 -2.6 -3.4 35 World* -3.7 -8.9 -2.7 -1.5 30 * Market exchange rate basis Source: Eikon Datastream 25 Exchange rates 2015 2016 2017 2018 2019 2020 2021 India Indonesia Japan end of period, # per US$ 2020 Dec-20 Jan-21 Feb-21 Source: Markit US$ broad index 112.1 112.1 111.8 112.3 • Economic conditions have been improving in Asia Japanese yen (JPY) 103.3 103.3 104.7 106.5 Pacific. Q4 GDP declines eased across most of the Australian dollar (AUD) 1.29 1.29 1.30 1.29 countries in the region. Moreover, as of February the Sth Korean won (KRW) 1087 1087 1114 1128 manufacturing PMI was back or above pre-crisis levels Indian rupee (INR) 73.1 73.1 73.0 73.5 across the three key markets that we regularly track. Indonesian rupiah (IDR) 14050 14050 14030 14240 Thai baht (THB) 30.0 30.0 29.9 30.1 • The trade-weighted US dollar index rose by 0.5%, Malaysian ringgit (MYR) 4.02 4.02 4.04 4.05 partly reflecting optimism about the US economic Source: Board of Governors of the Federal Reserve System (US), Eikon Datastream recovery. -

The Great Basin Landscape Conservation Cooperative
September 2010 The Great Basin Landscape Conservation Cooperative What is a Landscape Conservation The Great Basin Landscape Cooperative? Conservation Cooperative Landscape Conservation Cooperatives (LCCs) are applied science and management partnerships between Interior Department bureaus and others involved in natural resource management and What is the Great Basin LCC? conservation. Secretarial Order No. 3289, issued on Sept. 14, 2009 The Great Basin LCC will be a self-directed partnership. The by Interior Secretary Ken Salazar, calls for the establishment of 21 Great Basin LCC will provide a range of scientific and technical LCCs nationwide to better integrate science and management to support tools for landscape-scale conservation design to a wide address climate change and related issues array of managers. These tools will help managers identify and target biological objectives for native species and habitats in the What will the Great Basin LCC do? face of climate change and other stressors. Open public access to In broad terms, the Great Basin LCC will help link and integrate Great Basin LCC products will promote acceptance and use of Interior’s proposed Climate Science Centers with resource managers the science in regional conservation strategies. This effort is being and science users; will bring additional Interior resources to bear on coordinated with other regional partnerships and will be set up in landscape-scale issues and opportunities; and will help in applying a manner that will facilitate coordination and the identification science and facilitating coordination on a wide range of efforts of needs, capacities and gaps. We will link our efforts to bring to respond to climate change, invasive species, wildfires, human additional science capacity to improve conservation strategies development and other change agents across the Great Basin. -

Struggle for North America Prepare to Read
0120_wh09MODte_ch03s3_s.fm Page 120 Monday, June 4, 2007 10:26WH09MOD_se_CH03_S03_s.fm AM Page 120 Monday, April 9, 2007 10:44 AM Step-by-Step WITNESS HISTORY AUDIO SECTION 3 Instruction 3 A Piece of the Past In 1867, a Canadian farmer of English Objectives descent was cutting logs on his property As you teach this section, keep students with his fourteen-year-old son. As they focused on the following objectives to help used their oxen to pull away a large log, a them answer the Section Focus Question piece of turf came up to reveal a round, and master core content. 3 yellow object. The elaborately engraved 3 object they found, dated 1603, was an ■ Explain why the colony of New France astrolabe that had belonged to French grew slowly. explorer Samuel de Champlain. This ■ Analyze the establishment and growth astrolabe was a piece of the story of the of the English colonies. European exploration of Canada and the A statue of Samuel de Champlain French-British rivalry that followed. ■ Understand why Europeans competed holding up an astrolabe overlooks Focus Question How did European for power in North America and how the Ottawa River in Canada (right). their struggle affected Native Ameri- Champlain’s astrolabe appears struggles for power shape the North cans. above. American continent? Struggle for North America Prepare to Read Objectives In the 1600s, France, the Netherlands, England, and Sweden Build Background Knowledge L3 • Explain why the colony of New France grew joined Spain in settling North America. North America did not Given what they know about the ancient slowly. -

Countries and Continents of the World: a Visual Model
Countries and Continents of the World http://geology.com/world/world-map-clickable.gif By STF Members at The Crossroads School Africa Second largest continent on earth (30,065,000 Sq. Km) Most countries of any other continent Home to The Sahara, the largest desert in the world and The Nile, the longest river in the world The Sahara: covers 4,619,260 km2 The Nile: 6695 kilometers long There are over 1000 languages spoken in Africa http://www.ecdc-cari.org/countries/Africa_Map.gif North America Third largest continent on earth (24,256,000 Sq. Km) Composed of 23 countries Most North Americans speak French, Spanish, and English Only continent that has every kind of climate http://www.freeusandworldmaps.com/html/WorldRegions/WorldRegions.html Asia Largest continent in size and population (44,579,000 Sq. Km) Contains 47 countries Contains the world’s largest country, Russia, and the most populous country, China The Great Wall of China is the only man made structure that can be seen from space Home to Mt. Everest (on the border of Tibet and Nepal), the highest point on earth Mt. Everest is 29,028 ft. (8,848 m) tall http://craigwsmall.wordpress.com/2008/11/10/asia/ Europe Second smallest continent in the world (9,938,000 Sq. Km) Home to the smallest country (Vatican City State) There are no deserts in Europe Contains mineral resources: coal, petroleum, natural gas, copper, lead, and tin http://www.knowledgerush.com/wiki_image/b/bf/Europe-large.png Oceania/Australia Smallest continent on earth (7,687,000 Sq. -

The Diffusion of Maize to the Southwestern United States and Its Impact
PERSPECTIVE The diffusion of maize to the southwestern United States and its impact William L. Merrilla, Robert J. Hardb,1, Jonathan B. Mabryc, Gayle J. Fritzd, Karen R. Adamse, John R. Roneyf, and A. C. MacWilliamsg aDepartment of Anthropology, National Museum of Natural History, Smithsonian Institution, P.O. Box 37102, Washington, DC 20013-7012; bDepartment of Anthropology, One UTSA Circle, University of Texas at San Antonio, San Antonio, TX 78249; cHistoric Preservation Office, City of Tucson, P.O. Box 27210, Tucson, AZ 85726; dDepartment of Anthropology, Campus Box 1114, One Brookings Drive, Washington University, St. Louis, MO 63130; eCrow Canyon Archaeological Center, 23390 Road K, Cortez, CO 81321; fColinas Cultural Resource Consulting, 6100 North 4th Street, Private Mailbox #300, Albuquerque, NM 87107; and gDepartment of Archaeology, 2500 University Drive Northwest, University of Calgary, Calgary, Alberta, Canada T2N 1N4 Edited by Linda S. Cordell, University of Colorado, Boulder, CO, and approved October 30, 2009 (received for review June 22, 2009) Our understanding of the initial period of agriculture in the southwestern United States has been transformed by recent discoveries that establish the presence of maize there by 2100 cal. B.C. (calibrated calendrical years before the Christian era) and document the processes by which it was integrated into local foraging economies. Here we review archaeological, paleoecological, linguistic, and genetic data to evaluate the hypothesis that Proto-Uto-Aztecan (PUA) farmers migrating from a homeland in Mesoamerica intro- duced maize agriculture to the region. We conclude that this hypothesis is untenable and that the available data indicate instead a Great Basin homeland for the PUA, the breakup of this speech community into northern and southern divisions Ϸ6900 cal. -

Speleothem Paleoclimatology for the Caribbean, Central America, and North America
quaternary Review Speleothem Paleoclimatology for the Caribbean, Central America, and North America Jessica L. Oster 1,* , Sophie F. Warken 2,3 , Natasha Sekhon 4, Monica M. Arienzo 5 and Matthew Lachniet 6 1 Department of Earth and Environmental Sciences, Vanderbilt University, Nashville, TN 37240, USA 2 Department of Geosciences, University of Heidelberg, 69120 Heidelberg, Germany; [email protected] 3 Institute of Environmental Physics, University of Heidelberg, 69120 Heidelberg, Germany 4 Department of Geological Sciences, Jackson School of Geosciences, University of Texas, Austin, TX 78712, USA; [email protected] 5 Desert Research Institute, Reno, NV 89512, USA; [email protected] 6 Department of Geoscience, University of Nevada, Las Vegas, NV 89154, USA; [email protected] * Correspondence: [email protected] Received: 27 December 2018; Accepted: 21 January 2019; Published: 28 January 2019 Abstract: Speleothem oxygen isotope records from the Caribbean, Central, and North America reveal climatic controls that include orbital variation, deglacial forcing related to ocean circulation and ice sheet retreat, and the influence of local and remote sea surface temperature variations. Here, we review these records and the global climate teleconnections they suggest following the recent publication of the Speleothem Isotopes Synthesis and Analysis (SISAL) database. We find that low-latitude records generally reflect changes in precipitation, whereas higher latitude records are sensitive to temperature and moisture source variability. Tropical records suggest precipitation variability is forced by orbital precession and North Atlantic Ocean circulation driven changes in atmospheric convection on long timescales, and tropical sea surface temperature variations on short timescales. On millennial timescales, precipitation seasonality in southwestern North America is related to North Atlantic climate variability. -

Region: West Africa (14 Countries) (Benin, Burkina Faso, Cameroon, Chad, Côte D’Ivoire, Gambia, Ghana, Guinea, Guinea Bissau, Mali, Niger, Senegal, Togo)
Region: West Africa (14 Countries) (Benin, Burkina Faso, Cameroon, Chad, Côte d’Ivoire, Gambia, Ghana, Guinea, Guinea Bissau, Mali, Niger, Senegal, Togo) Project title: Emergency assistance for early detection and prevention of avian influenza in Western Africa Project number: TCP/RAF/3016 (E) Starting date: November 2005 Completion date: April 2007 Government counterpart Ministries of Agriculture responsible for project execution: FAO contribution: US$ 400 000 Signed: ..................................... Signed: ........................................ (on behalf of Government) Jacques Diouf Director-General (on behalf of FAO) Date of signature: ..................... Date of signature: ........................ I. BACKGROUND AND JUSTIFICATION In line with the FAO/World Organization for Animal Health (OIE) Global Strategy for the Progressive Control of Highly Pathogenic Avian Influenza (HPAI), this project has been developed to provide support to the regional grouping of West African countries to strengthen emergency preparedness against the eventuality of HPAI being introduced into this currently free area. There is growing evidence that the avian influenza, which has been responsible for serious disease outbreaks in poultry and humans in several Asian countries since 2003, is spread through a number of sources, including poor biosecurity at poultry farms, movement of poultry and poultry products and live market trade, illegal and legal trade in wild birds. Although unproven, it is also suspected that the virus could possibly be carried over long distances along the migratory bird flyways to regions previously unaffected (Table 1) is a cause of serious concern for the region. Avian influenza subtype H5N1 could be transported along these routes to densely populated areas in the South Asian Subcontinent and to the Middle East, Africa and Europe. -

The Northeast Region
The Northeast Region The Northeast Region Land and Water of the Northeast The region between the coast of the Atlantic Ocean and the Great Lakes is called the Northeast region. The Northeast region includes eleven states. The Northeast region has broad valleys, rolling hills, and low mountains. The Appalachian Mountains stretch from Maine in the Northeast region down to Alabama in the Southeast region. Several different mountain ranges make up the This map shows the eleven states of the Northeast Appalachian Mountains, including the region. Allegheny Mountains, the Pocono Mountains, the Adirondack Mountains, and the Catskills. The Appalachian Mountains are one of the oldest mountain ranges in the world. Although they used to be very tall, they are much smaller now. Over time, water, wind, and ice have caused the rock of the mountains to slowly break down. Now the peaks of the Appalachian Mountains are smooth and rounded. The longest hiking trail in the world is the Appalachian Trail. It goes from Maine to Georgia, along the spine of the mountain range, through more than 2,000 miles of valleys, hills, and rivers. The Appalachian Trail is a very popular place to hike. There are many people who have hiked the entire trail! The tops of the Appalachian Mountains have been worn down over many millions of years. ★ created by Jill S. Russ ★ mrsruss.com ★ Page 1 Mount Washington in New Hampshire is part of the Appalachian Mountain range. At 6,288 feet tall, Mount Washington is the highest peak in the Northeast. Mount Washington has some of the world's most severe weather. -

Americas COVID-19 Response Update #30
August 2021 Americas COVID-19 Response Update #30 “Over a year into the crisis, it is clear that COVID-19 has rolled back years of incremental gender equality and child protection gains and will continue to impact displaced women and children well into the future. Action is needed now to mitigate those impacts ” Gillian Triggs Assistant High Commissioner for Protection See the release of a data visualization tool on the negative impacts of COVID-19 on the displaces women and children HERE. Situational Highlights More than two million people in the increase during July. Guatemala, in As vaccination roll-out started across UNHCR and the Latin American Americas have died from COVID-19 particular, reported a high rate of the region, at least 22 countries have Network of Organizations of Persons complications, according to infections and hospitalizations, included refugees and asylum-see- with Disabilities and their Families WHO/PAHO, and less than 20% of mainly among people under 50. kers in their vaccination plan. (RIADIS) published a study on the people in Latin America and the challenges and resources for people Caribbean have been fully immuni- In the Caribbean, Cuba is experien- The countries in the region have with disabilities within the context of zed. cing higher COVID infection and relaxed their border restrictions over human mobility. Over 40% of people death rates than at any point during the past months. According to with disabilities consulted indicated As public health measures relax in the pandemic, aecting all age UNHCR’s monitoring on the impact that they have faced risks such as some countries, the number of cases groups. -

Regional Fact Sheet – North and Central America
SIXTH ASSESSMENT REPORT Working Group I – The Physical Science Basis Regional fact sheet – North and Central America Common regional changes • North and Central America (and the Caribbean) are projected to experience climate changes across all regions, with some common changes and others showing distinctive regional patterns that lead to unique combinations of adaptation and risk-management challenges. These shifts in North and Central American climate become more prominent with increasing greenhouse gas emissions and higher global warming levels. • Temperate change (mean and extremes) in observations in most regions is larger than the global mean and is attributed to human influence. Under all future scenarios and global warming levels, temperatures and extreme high temperatures are expected to continue to increase (virtually certain) with larger warming in northern subregions. • Relative sea level rise is projected to increase along most coasts (high confidence), and are associated with increased coastal flooding and erosion (also in observations). Exceptions include regions with strong coastal land uplift along the south coast of Alaska and Hudson Bay. • Ocean acidification (along coasts) and marine heatwaves (intensity and duration) are projected to increase (virtually certain and high confidence, respectively). • Strong declines in glaciers, permafrost, snow cover are observed and will continue in a warming world (high confidence), with the exception of snow in northern Arctic (see overleaf). • Tropical cyclones (with higher precipitation), severe storms, and dust storms are expected to become more extreme (Caribbean, US Gulf Coast, East Coast, Northern and Southern Central America) (medium confidence). Projected changes in seasonal (Dec–Feb, DJF, and Jun–Aug, JJA) mean temperature and precipitation at 1.5°C, 2°C, and 4°C (in rows) global warming relative to 1850–1900.