Common Mynas
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

FIRST CONTROL CAMPAIGN for COMMON MYNA (Acridotheres
FIRST CONTROL CAMPAIGN FOR COMMON MYNA ( Acridotheres tristis ) ON ASCENSION ISLAND 2009 By Susana Saavedra Project and field manager 1 Abstract This is a final report of the “First control campaign for Common myna (Acridotheres tristis ) in Ascension Island 2009”, which was undertaken as a private initiative of Live Arico Invasive Species Department. The field work took place, from the 25 th of September to 03 rd December 2009. Trapping was conducted in three phases: first on rubbish dumps and water tanks (29 days), second on a Sooty Tern Colony (15 days) and finally, again on rubbish dumps and water tanks (9 days). The goal of reducing the negative effects of the Common myna on native wildlife by trapping as many individuals as possible has been reasonably covered. The population of mynas, estimated in some 1.000 to 1.200 birds, has been reduced by culling 623 birds in 53 days. This work has been done by one person using four traps. There is a low risk of re-infestation from birds flying by their own from St Helena, and the only transport between Ascension and St Helena has been conveniently informed regarding mynas using boats as pathway and how to avoid it. Considering the present damage of the mynas for wildlife, human health and security, it is high recommended that the local Ascension Island Government or related Institutions should decide to go for eradication as soon as possible. Live Arico - P.O.Box 1132 38008 – Santa Cruz de Tenerife Canary Islands, Spain. 2 Live Arico Environmental and Animal Protection – Invasive Species Department Register Charity number: 4709 C.I.F.: G/ 38602058 e-mail : [email protected] Phone: + 34 620 126 525 I n d e x INTRODUCTION........................................................................................................... -

An Introduction to Birds and Birding
AN INTRODUCTION TO BIRDS AND BIRDWATCHING Illustration: Rohan Chakravarty Bird Count India www.birdcount.in [email protected] PART- I ABOUT INDIAN BIRDS From small to large Photos: Garima Bhatia / Rajiv Lather From common to rare Photos: Nirav Bhatt / Navendu Lad From nondescript to magnificent Photos: Mohanram Kemparaju / Rajiv Lather And from deserts to dense forests Photos: Clement Francis / Ramki Sreenivasan India is home to over 1200 species of birds! Photos: Dr. Asad Rahmani, Nikhil Devasar, Dhritiman Mukherjee, Ramana Athreya, Judd Patterson Birds in Indian Culture and Mythology Source: wikipedia.org Photo: Alex Loinaz Garuda, the vahana of Lord Vishnu is thought to be a Brahminy Kite Birds in Indian Culture and Mythology Source: wikipedia.org Jatayu, sacrificed himself to rescue Sita from being kidnapped by Ravana. He was thought to be a vulture. Birds in Indian Culture and Mythology Photo: Nayan Khanolkar Photo: Kalyan Varma Sarus Cranes have a strong cultural significance in North India for their fidelity while hornbills find mention in the traditional folklore of the tribes of Arunachal Pradesh. Bird behaviour: Foraging Illustration: Aranya Pathak Broome Bird behaviour: Foraging Photos: Mike Ross, Josep del Hoyo, Pat Bonish, Shreeram M.V Bird behaviour: Migration Photos: Arthur Morris / Dubi Shapiro | Maps: Greenland Institute of Natural Resources and R. Suresh Kumar Bird behaviour: Songs Recordings: Pronoy Baidya / Neils Poul Dreyer Threats to birds Photos: www.conservationindia.org Cartoon: Rohan Chakravarty What is -

How to Tell the Difference Between Noisy Miners and Indian Mynas
HOW TO TELL THE DIFFERENCE BETWEEN NOISY MINERS AND INDIAN MYNAS A lot of people want to know how to tell a noisy miner from an Indian myna. The Indian myna (also known as the common myna, common mynah or Indian mynah) has earned the reputation of being one of the worst feral animals in Australia. It’s likely that if you live in Sydney, Melbourne, North Queensland or Brisbane, you’re already familiar with it. This little brown bird might look harmless, but the World Conservation Union (IUCN) takes it very seriously. They put it on the list of the 100 most invasive species in the world and describe it as an extreme threat to Australia. Humans don’t get off easily either. Mynas carry bird mites and have the potential to carry avian-borne diseases that are dangerous to people, not to Noisy miner mention the huge amount of droppings they leave under their communal probably take the role of being the other birds to get the best nesting holes. roosting trees. Often gathering at night worst introduced species in Australia). in numbers more than a thousand, In the 1880s there was a locust You can discourage Indian mynas from these raucous birds can take over plague, so Indian mynas were brought your Australian garden by: clumps of trees, especially around in to control them. Of course, the • keeping less open grassed areas in areas where many people go (where mynas didn’t stop the locusts but your garden; they encounter fewer predators), such became another pest themselves. -

Species Threatenedsuperb Parrot Polytelis Swainsonii
Australian Species ThreatenedSuperb Parrot Polytelis swainsonii CONSERVATION STATUS COMMONWEALTH: Vulnerable (Environment Protection and Biodiversity Conservation Act 1999) AUSTRALIAN CAPITAL TERRITORY: Vulnerable (Nature Conservation Act 1980) NEW SOUTH WALES: Vulnerable (Threatened Species Conservation Act 1995) VICTORIA: Threatened (and listed under the Flora and Fauna Guarantee Act 1988) The Superb Parrot is a striking bird found in central woodland areas of southern WHERE DOES IT LIVE? New South Wales (NSW), the Australian The parrots are found in the NSW Capital Territory (ACT) and Victoria. southwest slopes as well as northern Already under threat from land clearing, parts of the ACT and north central loss of hollows, and lack of regeneration Victoria. Each spring they retreat towards of woodland habitat, this species may the southwest to breed, mainly in River soon be faced with another challenge and Blakely’s red gums. They then – the common myna bird. move further north and east, relying on Photo: Katherine Miller woodland habitat for flowers, fruits and seed, particularly in box and Blakely’s red gum. As one of the many Australian WHAT DOES IT bird species that uses tree hollows for breeding, clearing of woodland areas DID YOU KNOW... LOOK LIKE? has had a large impact on the parrot and, • The total population of the Superb The Superb Parrot is a medium-sized with minimal replacement of old trees, Parrot is estimated to be only a few bird with a long slender tail. Both males its numbers may continue to decline in thousand birds and females have a green body, although the future. • It is the official emblem of NSW’s the plumage on males tends to be more Boorowa Shire brilliant. -

New Amendment to Lacey Act Aids in Identification Of
Sater or Barrington - Int. 2879 I FISH AlJDWILDLIFESERVICE For Release JULY 9# 1954 NJ%!MENDMENT TO LACEXACT AIDS IN IDENTIFICATION OF SPECIESPROELBITED FOR IMPORT Aa an added safeguard against the introduction into the United States of aer- tain unwanted species of wild animals and birds, a new amendment to the Laoey Act has been adopted which clarifies the identification of these species, Secretary of the Interior Douglas McKay announced today. Among the provisions of the Lacey Act, passed on May 25, 1900, was one that prohibited the introduction into this country of certain species of wild animals and birds which might be detrimental to agriculture or horticultures In this provision of the a&, some of the species were actually nsmed, Since then, however, difficulty has bean experienced in making definite identifioation of these species because of the auceptance of revisionary work involving generic names, The new amendment, which specifies by scientific nsme those species that are to be excluded [except for certain specified uses and under rigidly controlled condi- tions mbodied in permits issued by the Fish and Wildlife Service), will not only aid persons planning to import oertain birds and animals, but will ease the work of port-of-entry authorit%es. As stated in the original act, prohibited species include: the mongoose (Herpeates a 0 tatu ); the so-called "flying-foxes" or fruit bats (all species of the Genus*T Ptero tiglish Sparrow (wer domestious); the starling (Sturnus vuJ.aar& ; "and such other birds and animals as the Secretary -

Jungle Myna (Acridotheres Fuscus)
Invasive animal risk assessment Biosecurity Queensland Agriculture Fisheries and Department of Jungle myna Acridotheres fuscus Steve Csurhes First published 2011 Updated 2016 © State of Queensland, 2016. The Queensland Government supports and encourages the dissemination and exchange of its information. The copyright in this publication is licensed under a Creative Commons Attribution 3.0 Australia (CC BY) licence. You must keep intact the copyright notice and attribute the State of Queensland as the source of the publication. Note: Some content in this publication may have different licence terms as indicated. For more information on this licence visit http://creativecommons.org/licenses/by/3.0/au/ deed.en" http://creativecommons.org/licenses/by/3.0/au/deed.en Front cover: Jungle myna Photo: Used with permission, Wikimedia Commons. Invasive animal risk assessment: Jungle myna Acridotheres fuscus 2 Contents Summary 4 Introduction 5 Identity and taxonomy 5 Description and biology 5 Diet 5 Reproduction 5 Preferred habitat and climate 6 Native range and global distribution 6 Current distribution and impact in Queensland 6 History as a pest overseas 7 Use 7 Potential distribution and impact in Queensland 7 References 8 Invasive animal risk assessment: Jungle myna Acridotheres fuscus 3 Summary Acridotheres fuscus (jungle myna) is native to an extensive area of India and parts of southeast Asia. Naturalised populations exist in Singapore, Taiwan, Fiji, Western Samoa and elsewhere. In Fiji, the species occasionally causes significant damage to crops of ground nuts, with crop losses of up to 40% recorded. Within its native range (South India), it is not a well documented pest, but occasionally causes considerable (localised) damage to fruit orchards. -

India: Kaziranga National Park Extension
INDIA: KAZIRANGA NATIONAL PARK EXTENSION FEBRUARY 22–27, 2019 The true star of this extension was the Indian One-horned Rhinoceros (Photo M. Valkenburg) LEADER: MACHIEL VALKENBURG LIST COMPILED BY: MACHIEL VALKENBURG VICTOR EMANUEL NATURE TOURS, INC. 2525 WALLINGWOOD DRIVE, SUITE 1003 AUSTIN, TEXAS 78746 WWW.VENTBIRD.COM INDIA: KAZIRANGA NATIONAL PARK EXTENSION February 22–27, 2019 By Machiel Valkenburg This wonderful Kaziranga extension was part of our amazing Maharajas’ Express train trip, starting in Mumbai and finishing in Delhi. We flew from Delhi to Guwahati, located in the far northeast of India. A long drive later through the hectic traffic of this enjoyable country, we arrived at our lodge in the evening. (Photo by tour participant Robert Warren) We enjoyed three full days of the wildlife and avifauna spectacles of the famous Kaziranga National Park. This park is one of the last easily accessible places to find the endangered Indian One-horned Rhinoceros together with a healthy population of Asian Elephant and Asiatic Wild Buffalo. We saw plenty individuals of all species; the rhino especially made an impression on all of us. It is such an impressive piece of evolution, a serious armored “tank”! On two mornings we loved the elephant rides provided by the park; on the back of these attractive animals we came very close to the rhinos. The fertile flood plains of the park consist of alluvial silts, exposed sandbars, and riverine flood-formed lakes called Beels. This open habitat is not only good for mammals but definitely a true gem for some great birds. Interesting but common birds included Bar-headed Goose, Red Junglefowl, Woolly-necked Stork, and Lesser Adjutant, while the endangered Greater Adjutant and Black-necked Stork were good hits in the stork section. -

Hand-Raising and Rehabilitation of Mynas
Hand-raising and rehabilitation of mynas Corina Gardner Hill mynas David Lim Introduction Mynas are average sized (about 22-28 cms) passerine birds which belong to the family of starlings, Sturnidea. The term ‘myna’ is commonly used to refer to starlings in India. Mynas are commonly distributed throughout Southern and Eastern Asia. These birds have duller plumage and are more terrestrial compared to other members of the starling family. 1 Rehabber’s Den © 2012 Hand-raising and rehabilitation of mynas Common myna Acridotheres tristis The common myna is widely distributed throughout India and Asia and has also been introduced to many parts of the world. The species lives in woodlands and near human habitations. They have brown plumage, a black head, throat and breast, while the bill and legs are yellow. They also have a distinctive yellow patch behind the eyes. They are omnivorous birds and will scavenge for just about anything including discarded scraps, insects, seeds, grain and fruit. They roost in large trees and build their nests in walls and rooftops of buildings. Common myna Tris Jungle myna Acridotheres fuscus Jungle mynas are found in and around the Indian subcontinent. They have brownish grey plumage, a tuft of feathers on their heads, white patches on their 2 Rehabber’s Den © 2012 Hand-raising and rehabilitation of mynas primaries and a white tipped tail. They typically live in forests, tea plantations and near paddy fields. They are omnivorous birds and their diet often consists of insects, fruit, seeds and even nectar from flowers. Jungle myna Devna Arora Bank myna Acridotheres ginginianus This species of myna is found primarily in the Indian subcontinent. -

Interspecific Communication Between the Common Myna (Acridotheres Tristis) and the White-Vented Myna (Acridotheres Grandis)
41 INTERSPECIFIC COMMUNICATION BETWEEN THE COMMON MYNA (ACRIDOTHERES TRISTIS) AND THE WHITE-VENTED MYNA (ACRIDOTHERES GRANDIS) Theerasak Jaimipak1, Somboon Kamtaeja2, Surakan Payakkhabutra3 & Supaluck Viruhpintu1,* ABSTRACT Sharing information about predators is a vital importance for birds that live in flocks. The Common myna (Acridotheres tristis) and the White-vented myna (Acridotheres grandis) usually live together in the same foraging and roosting flocks. Interspecific signals that could transfer crucial information between species need some similar structures for decoding the signals. Alarm and pre-flight calls of mynas were recorded when they were disturbed by human presence. Distress calls were recorded when the birds were captured from mixed-species foraging flocks. The structures of distress, alarm and pre-flight calls of mynas were analysed by using maximum frequency, minimum frequency, call length and bandwidth. Latent time of neck stretching and head turning behaviour were used to analyse the response of the birds. The results showed that the sonograms for distress, alarm and pre-flight calls for both species indicated high, medium and low levels of similarity. All sounds were interspecific communication, but alarm and pre-flight calls may carry more species specific elements than distress calls. This finding suggests that mynas could use distress, alarm and pre-flight calls to transfer crucial information in mixed-species flocks. Keywords: Common myna; White-vented myna; distress calls; alarm calls; pre-flight calls INTRODUCTION Animals get benefits from living with group members by receiving essential information. Groups of animals may be composed of different species that are able to share crucial information about resources or predators (Goodale et al., 2010). -

INDIAN MYNA BIRDS Acridotheres Tristis (Starling Family) | Common Names: Indian Myna, Common Myna
PEST CONTROL EST 1966 PEST FACT SHEET INDIAN MYNA BIRDS Acridotheres tristis (starling family) | Common names: Indian Myna, Common Myna Native habitat: Tropical Southern Asia from Iran to India and Sri Lanka However, the Indian Myna has spread to most of Southeast Asia, and parts of Oceania, North America, South Africa, and the Middle East. Like many Australian pests, the Indian Myna was unsuccessfully introduced into Melbourne in the 1860’s, and then later into Queensland, to keep the insect population down. The Indian Myna is brown with a black head, and has a yellow bill, legs, and bare eye skin. Not to be confused with the native Noisy Miner, which is mostly grey. CUREALLPEST.COM.AU (07) 3349 8572 PEST CONTROL EST 1966 PEST CONTROL EST 1966 PEST FACT SHEET INDIAN MYNA BIRDS Indian Mynas have been voted as Being an One of the World’s The Most Extreme 100 Most Hated Pest Threat Invasive Species in Australia National Vertebrate Pests Committee World Conservation Union ABC Wild Watch Quest for Pests 2005 But, why the hate? Well, Indian Mynas are: • Noisy, territorial, angry birds, and not afraid of humans. • During breeding season, they take over nests made by native birds • They kill the chicks and eggs of other birds • They build superfluous nests during the breeding season, eectively excluding native birds and animals from those nesting sites. CUREALLPEST.COM.AU (07) 3349 8572 PEST CONTROL EST 1966 PEST CONTROL EST 1966 PEST FACT SHEET INDIAN MYNA BIRDS So, what can you do? Here are some tips to keep Indian Mynas away: You could try to implement ...Or, you could just get help from these home-solutions.. -

The Distribution and Spread of the Invasive Alien Common Myna, Acridotheres Tristis L
Research Articles South African Journal of Science 103, November/December 2007 465 The distribution and spread of the invasive alien common myna, Acridotheres tristis L. (Aves: Sturnidae), in southern Africa Derick S. Peacocka, Berndt J. van Rensburga and Mark P. Robertsonb* result of human-induced habitat modification. The deteriora- The common myna is an Asian starling that has become established tion or fragmentation of habitats is not only conducive to alien in many parts of the world outside of its native range due to accidental establishment, but also works synergistically with the negative or deliberate introductions by humans. The South African population effects that the alien species themselves pose to remaining of this species originated from captive birds that escaped in Durban indigenous communities. in 1902. A century later, the common myna has become abundant Apart from the need to understand the processes and patterns throughout much of South Africa and is considered to pose a serious of biological invasions in order to minimize their adverse effects, threat to indigenous biodiversity. Preliminary observations suggest invasions are also of great academic interest because they provide that the common myna’s distribution is closely tied to that of quantifiable, relatively short-term examples of ecological and humans, but empirical evidence for this hypothesis is lacking. biogeographical processes. Owing to the large spatial and long We have investigated the relationships between common myna temporal scales required, experimental manipulations of popu- distribution, human population size and land-transformation values lations and communities for the investigation of, for example, at a quarter-degree resolution in South Africa. -
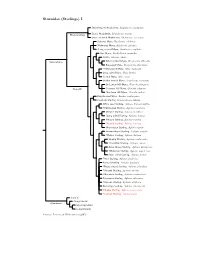
Sturnidae Tree, Part 1
Sturnidae (Starlings) I Stripe-headed Rhabdornis, Rhabdornis mystacalis Grand Rhabdornis, Rhabdornis grandis Rhabdornithini Stripe-breasted Rhabdornis, Rhabdornis inornatus Sulawesi Myna, Basilornis celebensis ?Helmeted Myna, Basilornis galeatus ?Long-crested Myna, Basilornis corythaix Apo Myna, Goodfellowia mirandus Coleto, Sarcops calvus Graculinae White-necked Myna, Streptocitta albicollis Bare-eyed Myna, Streptocitta albertinae ?Yellow-faced Myna, Mino dumontii Long-tailed Myna, Mino kreffti Golden Myna, Mino anais Golden-crested Myna, Ampeliceps coronatus Sri Lankan Hill-Myna, Gracula ptilogenys Graculini Common Hill Myna, Gracula religiosa ?Southern Hill Myna, Gracula indica Fiery-browed Myna, Enodes erythrophris Grosbeak Starling, Scissirostrum dubium White-eyed Starling, Aplonis brunneicapillus ?Yellow-eyed Starling, Aplonis mystacea Metallic Starling, Aplonis metallica ?Long-tailed Starling, Aplonis magna Pohnpei Starling, Aplonis pelzelni ?Kosrae Starling, Aplonis corvina Micronesian Starling, Aplonis opaca Brown-winged Starling, Aplonis grandis ?Makira Starling, Aplonis dichroa Singing Starling, Aplonis cantoroides ?Tanimbar Starling, Aplonis crassa Asian Glossy Starling, Aplonis panayensis ?Moluccan Starling, Aplonis mysolensis Short-tailed Starling, Aplonis minor ?Atoll Starling, Aplonis feadensis Rennell Starling, Aplonis insularis ?Rusty-winged Starling, Aplonis zelandica ?Striated Starling, Aplonis striata ?Mountain Starling, Aplonis santovestris Polynesian Starling, Aplonis tabuensis ?Samoan Starling, Aplonis atrifusca Rarotonga Starling, Aplonis cinerascens ?Mauke Starling, Aplonis mavornata ?Tasman Starling, Aplonis fusca Sturnini Cinnyricinclini Sturninae Onychognathini Lamprotornini Sources: Lovette and Rubenstein (2007)..