Avelines Hole
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Riding on Beaches and Estuaries
ADVICE ON Riding on Beaches and Estuaries 2 There are a number of beaches around England, Wales and Ireland that allow riding and BHS Approved centres that offer the opportunity to ride on a beach. There are many health benefits of riding on a sandy beach for horse and rider. Long sandy stretches are good for building up fitness levels and often the sand can encourage muscle tone and strength. It can provide outstanding views of the sea and is a refreshing way to see areas of beauty throughout the coasts of England, Wales and Ireland. Beach riding can be a wonderful experience for both you and your horse if you are aware of a few points of legality and safety, so please read all the guidance in this leaflet. Estuaries are where rivers meet the sea and they are unpredictable places, requiring caution and respect for the variety of conditions underfoot, the special ecology and the potential risks in riding there. While large expanses of open ground look inviting to riders, some of the conditions encountered may be dangerous. However, with due care and knowledge, estuaries can provide excellent riding opportunities. Is riding on the beach permitted? Check that riding on the beach is permitted. It may be limited to certain times, days or areas and there may be bylaws. Restrictions on time will often be to riders’ benefit, being at quieter periods such as early morning and late evening when there may be fewer other users to avoid. If there are areas where riding is not permitted, be sure you are clear about their extent and avoid them carefully; their boundaries may not be obvious even if they are above high water because signs and fences tend not to last long on the shore or may not be permitted. -
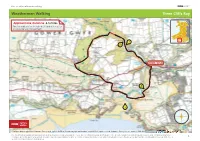
Weatherman Walking Three Cliffs Bay
bbc.co.uk/weathermanwalking © 2017 Weatherman Walking Three Cliffs Bay Approximate distance: 4.5 miles For this walk we’ve included OS grid references should you wish to use them. 1 2 Start End 4 3 N W E S Reproduced by permission of Ordnance Survey on behalf of HMSO. © Crown copyright and database right 2009.All rights reserved. Ordnance Survey Licence number 100019855 The Weatherman Walking maps are intended as a guide to help you walk the route. We recommend using an OS map of the area in conjunction with this guide. Routes and conditions may have changed since this guide was written. The BBC takes no responsibility for any accident or injury that may occur while following the route. Always wear appropriate clothing and footwear and check 1 weather conditions before heading out. bbc.co.uk/weathermanwalking © 2017 Weatherman Walking Three Cliffs Bay Start: Gower Heritage Centre, Parkmill Starting ref: SS 543 892 Distance: Approx. 4.5 miles Grade: Leisurely Walk time : 2 hours This delightful circular walk takes us through parkland, woodland, along a beach and up to an old castle high on a hill. Spectacular views abound and the sea air will ensure you sleep well at the end of it! We begin at the Gower Heritage Centre based around a working 12th century water mill where it’s worth spending some time fi nding out about the history of the area before setting off . Directions From the Heritage Centre, cross the ford then take the road to the right. Walk along for about a mile until you come to the entrance to Park Wood (Coed y Parc) on your right. -

Palaeolithic and Pleistocene Sites of the Mendip, Bath and Bristol Areas
Proc. Univ. Bristol Spelacol. Soc, 19SlJ, 18(3), 367-389 PALAEOLITHIC AND PLEISTOCENE SITES OF THE MENDIP, BATH AND BRISTOL AREAS RECENT BIBLIOGRAPHY by R. W. MANSFIELD and D. T. DONOVAN Lists of references lo works on the Palaeolithic and Pleistocene of the area were published in these Proceedings in 1954 (vol. 7, no. 1) and 1964 (vol. 10, no. 2). In 1977 (vol. 14, no. 3) these were reprinted, being then out of print, by Hawkins and Tratman who added a list ai' about sixty papers which had come out between 1964 and 1977. The present contribution is an attempt to bring the earlier lists up to date. The 1954 list was intended to include all work before that date, but was very incomplete, as evidenced by the number of older works cited in the later lists, including the present one. In particular, newspaper reports had not been previously included, but are useful for sites such as the Milton Hill (near Wells) bone Fissure, as are a number of references in serials such as the annual reports of the British Association and of the Wells Natural History and Archaeological Society, which are also now noted for the first time. The largest number of new references has been generated by Gough's Cave, Cheddar, which has produced important new material as well as new studies of finds from the older excavations. The original lists covered an area from what is now the northern limit of the County of Avon lo the southern slopes of the Mendips. Hawkins and Tratman extended that area to include the Quaternary Burtle Beds which lie in the Somerset Levels to the south of the Mendips, and these are also included in the present list. -

NLCA39 Gower - Page 1 of 11
National Landscape Character 31/03/2014 NLCA39 GOWER © Crown copyright and database rights 2013 Ordnance Survey 100019741 Penrhyn G ŵyr – Disgrifiad cryno Mae Penrhyn G ŵyr yn ymestyn i’r môr o ymyl gorllewinol ardal drefol ehangach Abertawe. Golyga ei ddaeareg fod ynddo amrywiaeth ysblennydd o olygfeydd o fewn ardal gymharol fechan, o olygfeydd carreg galch Pen Pyrrod, Three Cliffs Bay ac Oxwich Bay yng nglannau’r de i halwyndiroedd a thwyni tywod y gogledd. Mae trumiau tywodfaen yn nodweddu asgwrn cefn y penrhyn, gan gynnwys y man uchaf, Cefn Bryn: a cheir yno diroedd comin eang. Canlyniad y golygfeydd eithriadol a’r traethau tywodlyd, euraidd wrth droed y clogwyni yw bod yr ardal yn denu ymwelwyr yn eu miloedd. Gall y priffyrdd fod yn brysur, wrth i bobl heidio at y traethau mwyaf golygfaol. Mae pwysau twristiaeth wedi newid y cymeriad diwylliannol. Dyma’r AHNE gyntaf a ddynodwyd yn y Deyrnas Unedig ym 1956, ac y mae’r glannau wedi’u dynodi’n Arfordir Treftadaeth, hefyd. www.naturalresources.wales NLCA39 Gower - Page 1 of 11 Erys yr ardal yn un wledig iawn. Mae’r trumiau’n ffurfio cyfres o rostiroedd uchel, graddol, agored. Rheng y bryniau ceir tirwedd amaethyddol gymysg, yn amrywio o borfeydd bychain â gwrychoedd uchel i gaeau mwy, agored. Yn rhai mannau mae’r hen batrymau caeau lleiniog yn parhau, gyda thirwedd “Vile” Rhosili yn oroesiad eithriadol. Ar lannau mwy agored y gorllewin, ac ar dir uwch, mae traddodiad cloddiau pridd a charreg yn parhau, sy’n nodweddiadol o ardaloedd lle bo coed yn brin. Nodwedd hynod yw’r gyfres o ddyffrynnoedd bychain, serth, sy’n aml yn goediog, sydd â’u nentydd yn aberu ar hyd glannau’r de. -

2016 Directory the Dragon Hotel the Kingsway, Swansea SA1 5LS 01792 657100 [email protected] 4★H, 4★ H
2016 Directory The Dragon Hotel The Kingsway, Swansea SA1 5LS 01792 657100 www.dragon-hotel.co.uk [email protected] 4★H, 4★ H Serviced Accommodation Morgans Hotel Somerset Place, Swansea SA1 1RR 01792 484848 www.morganshotel.co.uk [email protected] 4★ H Self Catering Accommodation Caravan & Camping Somerfield Lodge B & B Clyne Golf Club, 118-120 Owls Lodge Lane, Mayals, Swansea SA3 5DP 01792 929293 www.somerfieldlodge.co.uk [email protected] 4★★GA Attractions & Activities Swansea Marriott Hotel Maritime Quarter, Swansea SA1 3SS 01792 642020 Food & Drink www.swanseamarriott.co.uk [email protected] 4★ H Travel Campus Accommodation, Swansea University Singleton Park, Swansea SA2 8PP 01792 295665 www.swansea.ac.uk/conferences [email protected] 3 - 4★ CA Abbreviations Accommodation Type Beachcomber 364 Oystermouth Road, Swansea SA1 3UL 01792 651380 AG Awaiting Grading AA Alternative Accommodation www.beachcomberguesthouse.com [email protected] 3★ GH APP Approved AcA Activity Accommodation BH Budget Hotel B Bunkhouse Mercure Swansea Phoenix Way, Swansea SA7 9EG 01792 310330 L Listed BB Bed & Breakfast www.mercure.com [email protected] 3★ H SP Service Provider CA Campus Accommodation ★ Visit Wales Grading F Farmhouse Village The Hotel Club Swansea Langdon Road, SA1 Waterfront, Swansea SA1 8QY 01792 341270 ★ AA grading GA Guest Accommodation www.village-hotels.com [email protected] Listed u AA grading for GpA Group Accommodation caravan parks -

Palaeolithic and Mesolithic
Palaeolithic and Mesolithic 2 Palaeolithic and Mesolithic Robert Hosfield, Vanessa Straker and Paula Gardiner with contributions from Anthony Brown, Paul Davies, Ralph Fyfe, Julie Jones and Heather Tinsley 2.1 Introduction For the Palaeolithic periods the open-landscape archaeology is dominated by lithic scatters (predom- The South West contains a diverse variety of Palaeo- inantly of deeply buried artefacts, frequently in fluvial lithic and Mesolithic archaeology of differing degrees deposits, and particularly true in the Lower and of significance. This reflects the nature of the arch- Middle Palaeolithic), although occupation sites such as aeological material itself, the histories of research Hengistbury Head (Barton 1992) and Kent’s Cavern in different parts of the region and, with regard to (Campbell and Sampson 1971) are also present. the Palaeolithic period, the differential preservation For the Mesolithic, there are greater numbers of of Pleistocene landforms and deposits throughout the excavated sites (especially from Somerset), although region. One of the key features of the Palaeolithic surface or shallow sub-surface lithic scatters are still archaeology is the presence of a significant cave-based common, especially in the west. resource in south Devon and northern Somerset, which is unquestionably of national significance (see Overall, the Palaeolithic and Mesolithic archae- for example, Campbell and Sampson 1971; Tratman ology of this region is generally rather poorly known, et al. 1971; Bishop 1975; Harrison 1977; Straw 1995; reflecting an absence of robust geochronological 1996; Andrews et al. 1999). frameworks, the predominance of research into a handful of cave and open sites over the lithic scatter In terms of an open-landscape Palaeolithic record, resource (whether located on the surface or deeply there is an inevitable bias towards those areas with buried) and the absence of any major syntheses. -

History Programme of Study
Historical Gower Key Stage 2 Park Wood Education Resource Notes for Teachers Contents Page Information for Teachers 1 How to use this pack 1 Risk Assessment 1 Equipment List 2 Curriculum Links 2 Cross - curricular work 3 Before you go activities 4 After your visit activities 4 Activities 1. Park Wood (Parc le Breos) 5 2. Giants Grave 6 3. Cathole Cave 7 4. Park Wood limekiln and quarries 8 Park Wood trail leaflet 9 Credits This education pack was written and designed by Audio Trails Ltd (www.audiotrails.co.uk) on behalf of Gower Landscape Partnership. The Gower Landscape Project has received funding through the Rural Development Plan for Wales 2007-2013, which is funded by the Welsh Government and the European Agricultural Fund for Rural Development, and also from the Heritage Lottery Fund under its Landscape Partnership programme. Other funding partners include the City & County of Swansea, Natural Resources Wales and The National Trust. Images were supplied and are copyright of the following individuals and organisations: Audio Trails Ltd © Copyright GGAT HER Charitable Trust Helen Grey Information for teachers Gower’s rich varied landscape has made it an attractive environment for human occupation since at least c 123,000 BC. The peninsula has been home to Stone Age hunter-gatherers, Iron Age farmers and warriors, early Christian immigrants and Norman knights. This part of the Gower app around Park Wood explores the theme ‘Historical Gower’. The trail has four stops: Giants Grave, Cathole Cave, Limekiln and quarries and Parc le Breos. At each stop oral histories, stories, photographs, facts and information are used to reveal the areas fascinating historical past. -

Ancient Genomes Indicate Population Replacement in Early Neolithic Britain
SUPPLEMENTARY INFORMATIONARTICLES https://doi.org/10.1038/s41559-019-0871-9 In the format provided by the authors and unedited. Ancient genomes indicate population replacement in Early Neolithic Britain Selina Brace1,15, Yoan Diekmann2,15, Thomas J. Booth1,15, Lucy van Dorp 3, Zuzana Faltyskova2, Nadin Rohland4, Swapan Mallick3,5,6, Iñigo Olalde4, Matthew Ferry4,6, Megan Michel4,6, Jonas Oppenheimer4,6, Nasreen Broomandkhoshbacht4,6, Kristin Stewardson4,6, Rui Martiniano 7, Susan Walsh8, Manfred Kayser 9, Sophy Charlton 1,10, Garrett Hellenthal3, Ian Armit 11, Rick Schulting12, Oliver E. Craig 10, Alison Sheridan13, Mike Parker Pearson14, Chris Stringer 1, David Reich4,5,6,16, Mark G. Thomas 2,3,16* and Ian Barnes 1,16* 1Department of Earth Sciences, Natural History Museum, London, UK. 2Research Department of Genetics, Evolution and Environment, University College London, London, UK. 3UCL Genetics Institute, University College London, London, UK. 4Department of Genetics, Harvard Medical School, Boston, MA, USA. 5Broad Institute of MIT and Harvard, Cambridge, MA, USA. 6Howard Hughes Medical Institute, Harvard Medical School, Boston, MA, USA. 7Department of Genetics, University of Cambridge, Cambridge, UK. 8Department of Biology, Indiana University-Purdue University Indianapolis, Indianapolis, IN, USA. 9Department of Genetic Identification, Erasmus University Medical Centre Rotterdam, Rotterdam, the Netherlands. 10Bioarch, University of York, York, UK. 11School of Archaeological and Forensic Sciences, University of Bradford, Bradford, UK. 12Institute of Archaeology, University of Oxford, Oxford, UK. 13National Museums Scotland, Edinburgh, UK. 14Institute of Archaeology, University College London, London, UK. 15These authors contributed equally: Selina Brace, Yoan Diekmann, Thomas J. Booth. 16These authors jointly supervised this work: David Reich, Mark G. -

Programme – Swansea Ramblers We Offer Enjoyable Short & Long Walks
Programme – Swansea Ramblers We offer enjoyable short & long walks all year around and welcome new walkers to try a walk with us. 1 Front Cover Photograph: A Ramblers’ visit to Llanmadoc Church v23 2 About Swansea Ramblers Swansea Ramblers, (originally West Glamorgan Ramblers) was formed in 1981. We always welcome new walkers to share our enjoyment of the countryside, socialise and make new friends. We organise long and short walks, varying from easy to strenuous across a wide area of South and Mid Wales, including Gower and Swansea. Swansea Ramblers Website: www.swansearamblers.org.uk On the website, you’ll find lots of interest and photographs of previous walks. For many new members, this is their first introduction to our group and part of the reason they choose to walk with us. Programme of walks: We have walks to suit most tastes. The summer programme runs from April to September and the winter programme covers October to March. A copy of the programme is supplied to members and can be downloaded from our website. Evening short walks: These are about 2-3 miles and we normally provide these popular walks once a week in the summer. Monday Short walks: These are 2-3 mile easier walks as an introduction to walking and prove popular with new walkers. Weekday walks: We have one midweek walk each week. The distance can vary from week to week, as can the day on which it takes place. Saturday walks: We have a Saturday walk every week that is no more than 6 miles in length and these are a great way to begin exploring the countryside. -

Excavations at Cathole Cave, Gower, Swansea
Proc. Univ. Bristol Spelaeol. Soc., 2014, 26 (2), 131-169 EXCAVATIONS AT CATHOLE CAVE, GOWER, SWANSEA by ELIZABETH A. WALKER, DAVID CASE, CLAIRE INGREM, JENNIFER R. JONES and RICHARD MOURNE ABSTRACT The discovery of an engraving in Cathole Cave in 2010 led to a decision to grille the cave. In 2012 excava- tions took place in the cave ahead of the grilling. Two areas of the cave were excavated; Trench A demonstrating that the cave held a faunal occupation dating to MIS 3, at a time, or times, between 50,000 and 30,000 BP. Two flint blades of Upper Palaeolithic appearance were discovered along with a faunal assemblage from within the shallow deposits across the line of the grille. The work demonstrated that the earlier excavations in the cave by Col. E.R. Wood were extensive and at this point in the cave he excavated to bedrock. The standing section (Area B) which he left exposed further along the main cave passage was also cleaned, recorded and sampled. The deposits contain a faunal assemblage dominated by microfauna, but no cultural artefacts were found. The dating of key animal bones, the analysis of the microfauna and the sedimentological analysis have together enabled a picture to be developed of the changes in this section from the mid-Devensian to Late Glacial and Holocene. INTRODUCTION Cathole Cave is located in an inland dry valley, Green Cwm, Gower, Swansea, (NGR: SS 53764 90011; Figure 1). The cave lies in a fractured and weathered limestone cliff above the valley in which the Neolithic Severn Cotswold tomb of Parc le Breos Cwm is sited. -
Legendary Adventures
Play Safe Finding Out visitswanseabay.com Whatever you do and wherever you do it, please Visitor Information Points ensure that the conditions are safe and that you Where you see this sign, our friendly VIP’s have the right equipment. If new to the activity or can help you with: inexperienced, use an accredited operator (where • Ideas on where to go and things to do appropriate, all operators featured in this Guide • Finding local accommodation Legendary have achieved the relevant activity accreditation • Travel information in relation to the sections in which they appear). To help you find your nearest VIP, go to If participating in an activity in the sea or near the visitswanseabay.com/vips shoreline, then keep an eye on the state of the tide, Adventures as it can turn very quickly here, especially at the Worm’s Head causeway, Burry Holms and If you require any additional information regarding Three Cliffs Bay. accommodation, events and things to do, then visit: visitswanseabay.com If available, always follow warning flags and notices. Lifeguard patrolled beaches are best for swimming, Visit our Facebook,Twitter and Instagram pages especially for children. If surfing at Llangennith for more inspiration and local updates. watch out for shipwrecks below the waterline. Use @visitswanseabay and #LiveTheLegend when you post and we can join in your RNLI Legendary Swansea Bay Adventure! rnli.org WALES’ PREMIER For more information on dog friendly beaches, LEISURE DESTINATION accommodation and attractions Spend some quality time with the family! go to: Spend some quality time visitswanseabay.com/ dog-friendly-holidays Getting About with the family! Call our pre-booking hotline Swansea Bay is at the heart of South Wales and on 01792 466500 now! can be easily reached by motorway, with express This publication is available coaches and fast train links. -

Year 3, Spring: Stone Age to Iron Age History What Should I Already Know?
St Paul’s Church of England VC Junior School Knowledge Organiser Year 3, Spring: Stone age to Iron Age History What should I already know? Skills to be taught An awareness of the past and the passing of Understand that pre-history occurred before historical records were kept. time. Understand that prehistorical periods (Palaeolithic, Mesolithic, Neolithic) occurred over many thousands of years. Identify similarities and differences between Analyse and sequence changes in human’s existence from hunter/gatherer, settlement building to beginnings of different ways of life in different periods. agriculture. Understand how to find out about the past and Compare stone age existence with our life today. different ways of representing that. Explain how changes over time made an impact on the way humans survived and developed. Select and record information for study, begin to use library and internet resources to build upon knowledge. What will I know by the end of the unit? Vocabulary There were 3 distinct periods of time Archaeologist Person who learns about the past through digging up artefacts and studying them. during which significant developments in artefact An object made by a human being. human existence occurred. Palaeolithic: humans were hunter bronze An alloy or mixture of tin and copper. gatherers who led a nomadic existence, Bronze age A period of time between the Stone and Iron Ages. In Britain, the Bronze Age started around 2100BC moving with the seasons. Tools began and ended around 650BC. being fashioned (hand axe) Cheddar Cave Cheddar caves is where Britain's oldest complete human skeleton was found.