Social Ecology and Behavior of Coyotes
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Longest Terrestrial Migrations and Movements Around the World Kyle Joly 1*, Eliezer Gurarie2, Mathew S
www.nature.com/scientificreports OPEN Longest terrestrial migrations and movements around the world Kyle Joly 1*, Eliezer Gurarie2, Mathew S. Sorum3, Petra Kaczensky 4,5, Matthew D. Cameron 1, Andrew F. Jakes6, Bridget L. Borg7, Dejid Nandintsetseg8,9, J. Grant C. Hopcraft10, Bayarbaatar Buuveibaatar11, Paul F. Jones12, Thomas Mueller8,9, Chris Walzer 5,13, Kirk A. Olson11, John C. Payne5,11,13, Adiya Yadamsuren14,15 & Mark Hebblewhite16 Long-distance terrestrial migrations are imperiled globally. We determined both round-trip migration distances (straight-line measurements between migratory end points) and total annual movement (sum of the distances between successive relocations over a year) for a suite of large mammals that had potential for long-distance movements to test which species displayed the longest of both. We found that caribou likely do exhibit the longest terrestrial migrations on the planet, but, over the course of a year, gray wolves move the most. Our results were consistent with the trophic-level based hypothesis that predators would move more than their prey. Herbivores in low productivity environments moved more than herbivores in more productive habitats. We also found that larger members of the same guild moved less than smaller members, supporting the ‘gastro-centric’ hypothesis. A better understanding of migration and movements of large mammals should aid in their conservation by helping delineate conservation area boundaries and determine priority corridors for protection to preserve connectivity. The magnitude of the migrations and movements we documented should also provide guidance on the scale of conservation eforts required and assist conservation planning across agency and even national boundaries. -

Plant Species of Special Concern and Vascular Plant Flora of the National
Plant Species of Special Concern and Vascular Plant Flora of the National Elk Refuge Prepared for the US Fish and Wildlife Service National Elk Refuge By Walter Fertig Wyoming Natural Diversity Database The Nature Conservancy 1604 Grand Avenue Laramie, WY 82070 February 28, 1998 Acknowledgements I would like to thank the following individuals for their assistance with this project: Jim Ozenberger, ecologist with the Jackson Ranger District of Bridger-Teton National Forest, for guiding me in his canoe on Flat Creek and for providing aerial photographs and lodging; Jennifer Whipple, Yellowstone National Park botanist, for field assistance and help with field identification of rare Carex species; Dr. David Cooper of Colorado State University, for sharing field information from his 1994 studies; Dr. Ron Hartman and Ernie Nelson of the Rocky Mountain Herbarium, for providing access to unmounted collections by Michele Potkin and others from the National Elk Refuge; Dr. Anton Reznicek of the University of Michigan, for confirming the identification of several problematic Carex specimens; Dr. Robert Dorn for confirming the identification of several vegetative Salix specimens; and lastly Bruce Smith and the staff of the National Elk Refuge for providing funding and logistical support and for allowing me free rein to roam the refuge for plants. 2 Table of Contents Page Introduction . 6 Study Area . 6 Methods . 8 Results . 10 Vascular Plant Flora of the National Elk Refuge . 10 Plant Species of Special Concern . 10 Species Summaries . 23 Aster borealis . 24 Astragalus terminalis . 26 Carex buxbaumii . 28 Carex parryana var. parryana . 30 Carex sartwellii . 32 Carex scirpoidea var. scirpiformis . -

The Evolution of Micro-Cursoriality in Mammals
© 2014. Published by The Company of Biologists Ltd | The Journal of Experimental Biology (2014) 217, 1316-1325 doi:10.1242/jeb.095737 RESEARCH ARTICLE The evolution of micro-cursoriality in mammals Barry G. Lovegrove* and Metobor O. Mowoe* ABSTRACT Perissodactyla) in response to the emergence of open landscapes and In this study we report on the evolution of micro-cursoriality, a unique grasslands following the Eocene Thermal Maximum (Janis, 1993; case of cursoriality in mammals smaller than 1 kg. We obtained new Janis and Wilhelm, 1993; Yuanqing et al., 2007; Jardine et al., 2012; running speed and limb morphology data for two species of elephant- Lovegrove, 2012b; Lovegrove and Mowoe, 2013). shrews (Elephantulus spp., Macroscelidae) from Namaqualand, Loosely defined, cursorial mammals are those that run fast. South Africa, which we compared with published data for other However, more explicit definitions of cursoriality remain obscure mammals. Elephantulus maximum running speeds were higher than because locomotor performance is influenced by multiple variables, those of most mammals smaller than 1 kg. Elephantulus also including behaviour, biomechanics, physiology and morphology possess exceptionally high metatarsal:femur ratios (1.07) that are (Taylor et al., 1970; Garland, 1983a; Garland, 1983b; Garland and typically associated with fast unguligrade cursors. Cursoriality evolved Janis, 1993; Stein and Casinos, 1997; Carrano, 1999). In an in the Artiodactyla, Perissodactyla and Carnivora coincident with evaluation of these definition problems, Carrano (Carrano, 1999) global cooling and the replacement of forests with open landscapes argued that ‘…morphology should remain the fundamental basis for in the Oligocene and Miocene. The majority of mammal species, making distinctions between locomotor performance…’. -

How to Design Experiments in Animal Behaviour∗ 8
SERIES ARTICLE How to Design Experiments in Animal Behaviour∗ 8. How Do Wasps Decide Who Would Be the Queen? Part 2 Raghavendra Gadagkar Continuing to explore the fascinating world of the Indian pa- per wasp Ropalidia marginata, in this article, we will ask how wasps choose their queens in another context. In the previ- ous article in this series, we saw how a simple experiment re- vealed that wasps fight, i.e., indulge in dominance-subordinate interactions, and the winner becomes the queen and the loser becomes the worker. This was in the context of new nestfoundation. But contextmatters. Whenthe same wasps once again have to decide who will be their next queen Raghavendra Gadagkar is if the first one dies or is experimentally removed, the same DST Year of Science Chair Professor at the Centre for rules do not hold. The wasps in a mature colony continue Ecological Sciences, Indian to show dominance-subordinate interactions and can even be Institute of Science, arranged in a dominance hierarchy, but the dominance ranks Bangalore, Honorary of the wasps do not predict who their next queen will be. Professor at JNCASR, and Non-Resident Permanent How they choose their next queen in this context continues Fellow of the to be an enduring mystery. In this article, I will describe four Wissenschaftskolleg (Institute simple experiments that have helped us come close to nail- for Advanced Study), Berlin. ing the culprit, although I must confess that we have not yet During the past 40 years he has established an active found the smoking gun—the chase is on, and we are hot on school of research in the area the trail—please join in! of animal behaviour, ecology and evolution. -

Comparative Methods Offer Powerful Insights Into Social Evolution in Bees Sarah Kocher, Robert Paxton
Comparative methods offer powerful insights into social evolution in bees Sarah Kocher, Robert Paxton To cite this version: Sarah Kocher, Robert Paxton. Comparative methods offer powerful insights into social evolution in bees. Apidologie, Springer Verlag, 2014, 45 (3), pp.289-305. 10.1007/s13592-014-0268-3. hal- 01234748 HAL Id: hal-01234748 https://hal.archives-ouvertes.fr/hal-01234748 Submitted on 27 Nov 2015 HAL is a multi-disciplinary open access L’archive ouverte pluridisciplinaire HAL, est archive for the deposit and dissemination of sci- destinée au dépôt et à la diffusion de documents entific research documents, whether they are pub- scientifiques de niveau recherche, publiés ou non, lished or not. The documents may come from émanant des établissements d’enseignement et de teaching and research institutions in France or recherche français ou étrangers, des laboratoires abroad, or from public or private research centers. publics ou privés. Apidologie (2014) 45:289–305 Review article * INRA, DIB and Springer-Verlag France, 2014 DOI: 10.1007/s13592-014-0268-3 Comparative methods offer powerful insights into social evolution in bees 1 2 Sarah D. KOCHER , Robert J. PAXTON 1Department of Organismic and Evolutionary Biology, Museum of Comparative Zoology, Harvard University, Cambridge, MA, USA 2Institute for Biology, Martin-Luther-University Halle-Wittenberg, Halle, Germany Received 9 September 2013 – Revised 8 December 2013 – Accepted 2 January 2014 Abstract – Bees are excellent models for studying the evolution of sociality. While most species are solitary, many form social groups. The most complex form of social behavior, eusociality, has arisen independently four times within the bees. -

Behavioural Plasticity and the Transition to Order in Jackdaw Flocks
ARTICLE https://doi.org/10.1038/s41467-019-13281-4 OPEN Behavioural plasticity and the transition to order in jackdaw flocks Hangjian Ling 1,2, Guillam E. Mclvor 3, Joseph Westley3, Kasper van der Vaart1, Richard T. Vaughan4, Alex Thornton 3* & Nicholas T. Ouellette 1* Collective behaviour is typically thought to arise from individuals following fixed interaction rules. The possibility that interaction rules may change under different circumstances has 1234567890():,; thus only rarely been investigated. Here we show that local interactions in flocks of wild jackdaws (Corvus monedula) vary drastically in different contexts, leading to distinct group- level properties. Jackdaws interact with a fixed number of neighbours (topological interac- tions) when traveling to roosts, but coordinate with neighbours based on spatial distance (metric interactions) during collective anti-predator mobbing events. Consequently, mobbing flocks exhibit a dramatic transition from disordered aggregations to ordered motion as group density increases, unlike transit flocks where order is independent of density. The relationship between group density and group order during this transition agrees well with a generic self- propelled particle model. Our results demonstrate plasticity in local interaction rules and have implications for both natural and artificial collective systems. 1 Department of Civil and Environmental Engineering, Stanford University, Stanford, CA 94305, USA. 2 Department of Mechanical Engineering, University of Massachusetts Dartmouth, North Dartmouth, -

Techniques for Estimating Coyote Abundance
University of Nebraska - Lincoln DigitalCommons@University of Nebraska - Lincoln Symposium Proceedings—Coyotes in the Southwest: A Compendium of Our Knowledge Wildlife Damage Management, Internet Center (1995) for April 1995 TECHNIQUES FOR ESTIMATING COYOTE ABUNDANCE Scott E. Henke Texas A&M University- Kingsville, Kingsville, TX Fred F. Knowlton USDA-APHIS Follow this and additional works at: https://digitalcommons.unl.edu/coyotesw Part of the Environmental Health and Protection Commons Henke, Scott E. and Knowlton, Fred F., "TECHNIQUES FOR ESTIMATING COYOTE ABUNDANCE" (1995). Symposium Proceedings—Coyotes in the Southwest: A Compendium of Our Knowledge (1995). 28. https://digitalcommons.unl.edu/coyotesw/28 This Article is brought to you for free and open access by the Wildlife Damage Management, Internet Center for at DigitalCommons@University of Nebraska - Lincoln. It has been accepted for inclusion in Symposium Proceedings—Coyotes in the Southwest: A Compendium of Our Knowledge (1995) by an authorized administrator of DigitalCommons@University of Nebraska - Lincoln. TECHNIQUES FOR ESTIMATING COYOTE ABUNDANCE SCOTT E. HENKE, Caesar Kleberg Wlldlife Research Institute, Campus Box 2 18, Texas A&M University- Kingsvllle, Kingsville, TX 78363 FRED F. KNOWLTON, USDA-APHIS Denver Wildlife Research Center, Utah State Univers~ty,Logan, Utah 84322-5295 Absirad Knowledge of coyote abundance is needed to make intelligent management decisions Several methods have been devised to ennumerate coyote (Canis latrans) population size. We review several techniques and attempt to identify biases associated with each method. Once biases are understood, recommendations can be made to minimize theu impact on data collection processes and yield better estimates of coyote population trends. Enumerat~onof population status (i e ,denslty, Methods used to estimate coyote population trends) is impostant in research and management of slze, dens~ty,and relative abundance have included wildlife. -

Wildlife Ecology Provincial Resources
MANITOBA ENVIROTHON WILDLIFE ECOLOGY PROVINCIAL RESOURCES !1 ACKNOWLEDGEMENTS We would like to thank: Olwyn Friesen (PhD Ecology) for compiling, writing, and editing this document. Subject Experts and Editors: Barbara Fuller (Project Editor, Chair of Test Writing and Education Committee) Lindsey Andronak (Soils, Research Technician, Agriculture and Agri-Food Canada) Jennifer Corvino (Wildlife Ecology, Senior Park Interpreter, Spruce Woods Provincial Park) Cary Hamel (Plant Ecology, Director of Conservation, Nature Conservancy Canada) Lee Hrenchuk (Aquatic Ecology, Biologist, IISD Experimental Lakes Area) Justin Reid (Integrated Watershed Management, Manager, La Salle Redboine Conservation District) Jacqueline Monteith (Climate Change in the North, Science Consultant, Frontier School Division) SPONSORS !2 Introduction to wildlife ...................................................................................7 Ecology ....................................................................................................................7 Habitat ...................................................................................................................................8 Carrying capacity.................................................................................................................... 9 Population dynamics ..............................................................................................................10 Basic groups of wildlife ................................................................................11 -
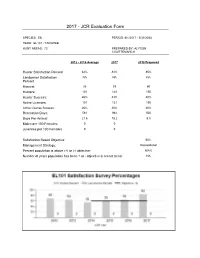
2017 - JCR Evaluation Form
2017 - JCR Evaluation Form SPECIES: Elk PERIOD: 6/1/2017 - 5/31/2018 HERD: EL101 - TARGHEE HUNT AREAS: 73 PREPARED BY: ALYSON COURTEMANCH 2012 - 2016 Average 2017 2018 Proposed Hunter Satisfaction Percent 62% 84% 85% Landowner Satisfaction NA NA NA Percent Harvest: 26 53 60 Hunters: 101 124 150 Hunter Success: 26% 43% 40% Active Licenses: 101 131 150 Active License Success: 26% 40% 40% Recreation Days: 561 964 500 Days Per Animal: 21.6 18.2 8.3 Males per 100 Females: 0 0 Juveniles per 100 Females 0 0 Satisfaction Based Objective 60% Management Strategy: Recreational Percent population is above (+) or (-) objective: N/A% Number of years population has been + or - objective in recent trend: NA 2018 HUNTING SEASONS TARGHEE ELK HERD (EL101) Hunt Season Dates Type Quota License Limitations Area Opens Closes 73 Sep. 20 Oct. 25 General Antlered elk, spikes excluded 6 Aug. 15 Jan. 31 25 Limited Cow or calf valid on private quota land Archery Sep. 1 Sep.19 Refer to Section 3 of this Chapter Management Evaluation Current Hunter/Landowner Satisfaction Management Objective: Primary Objective: Achieve a 3-year average of ≥ 60% of hunters indicating they are “satisfied” or “very satisfied” on the harvest survey. Secondary Objective: Achieve a 3-year average of ≥ 25% harvest success. Management Strategy: Recreational 2017 Hunter Satisfaction Estimate: 84% 2017 Hunter Success: 43% Most Recent 3-Year Running Average Hunter Satisfaction Estimate: 69% Most Recent 3-Year Running Average Hunter Success: 31% The Wyoming Game and Fish Department (WGFD) proposed changing the objective for the Targhee Elk Herd from a postseason population objective to a hunter satisfaction objective in 2014. -

Eusociality: Origin and Consequences
Corrections CELL BIOLOGY. For the article ‘‘The NF1 tumor suppressor criti- BIOCHEMISTRY. For the article ‘‘Engineered single-chain dimeric cally regulates TSC2 and mTOR,’’ by Cory M. Johannessen, streptavidins with an unexpected strong preference for biotin- Elizabeth E. Reczek, Marianne F. James, Hilde Brems, Eric 4-fluorescein,’’ by Filiz M. Aslan, Yong Yu, Scott C. Mohr, and Legius, and Karen Cichowski, which appeared in issue 24, June Charles R. Cantor, which appeared in issue 24, June 14, 2005, of 14, 2005, of Proc. Natl. Acad. Sci. USA (102, 8573–8578; first Proc. Natl. Acad. Sci. USA (102, 8507–8512; first published June published June 3, 2005; 10.1073͞pnas.0503224102), the authors 6, 2005; 10.1073͞pnas.0503112102), the footnotes appeared in note the following. On page 8574, the last sentence of the first the wrong order, due to a printer’s error. The ** footnote on paragraph in the left column, ‘‘Clarified lysates were normalized page 8507 should have read: ‘‘Throughout this paper, we use the for protein levels and analyzed by Western blotting with the terms monomer(ic), dimer(ic), and tetramer(ic) to refer to the following antibodies: phospho-p70S6K (T-389), phospho- number of biotin-binding domains present in a molecule, re- tuberin (S-939), phospho-tuberin (T-1462), phospho-Akt (S- gardless of the number of polypeptide chains it has.’’ In addition, 473), tuberin (C-20) from Santa Cruz Biotechnology, and the †† footnote on page 8512 should have read: ‘‘We note, protein kinase B␣͞AKT1, actin, and -tubulin from Sigma- however, -

Finding a Mate with Eusocial Skills
Finding a Mate With Eusocial Skills Chris Marriott1 and Jobran Chebib2 1University of Washington, Tacoma, WA, USA 98402 2University of Zurich,¨ Zurich,¨ Switzerland 8057 [email protected] Downloaded from http://direct.mit.edu/isal/proceedings-pdf/alif2016/28/298/1904330/978-0-262-33936-0-ch052.pdf by guest on 30 September 2021 Abstract mutual response of the individuals in the herd with no need for social awareness or exchange of information. Sexual reproductive behavior has a necessary social coordina- Prior work (from now on when we reference the prior tion component as willing and capable partners must both be work we mean the work in Marriott and Chebib (2015a,b)) in the right place at the right time. It has recently been demon- strated that many social organizations that support sexual re- showed that herding, philopatry, and assortative mating production can evolve in the absence of social coordination arose through non-social mechanisms of convergence and between agents (e.g. herding, assortative mating, and natal common descent. This work raised a few questions regard- philopatry). In this paper we explore these results by includ- ing the role that social interaction plays in many of these ob- ing social transfer mechanisms to our agents and contrasting served behaviors. These mating behaviors can be explained their reproductive behavior with a control group without so- cial transfer mechanisms. We conclude that similar behaviors by both non-social and social mechanisms. In many cases emerge in our social learning agents as those that emerged in the non-social solution is the simpler one, though it is likely the non-social learning agents. -

Close Cache Creek for Cougars Records Show Game Tain Lions Using the Area,” Fuchs Con- Tinued
Black JACKSON HOLE NEWS&GUIDE, Wednesday, February 22, 2012 - 9A Residents: Close Cache Creek for cougars Records show Game tain lions using the area,” Fuchs con- tinued. “On Thursday night, however, and Fish inundated the family group returned and killed with comments after another deer. At that point the prop- erty owner asked if there was more cat family was captured. that we could do to haze her and the siblings from their property.” By Cory Hatch In the email, Fuchs expressed a de- sire to keep the family group intact. Some Jackson Hole residents said “It is our hope that we will be able they would rather close Cache Creek to capture at least the female quick- temporarily than see a cougar fam- ly,” he said. “If so, we will attempt to ily relocated from its home in the tree and immobilize the kittens if we drainage. cannot capture them in the box traps. Over several weeks, wildlife man- COURTESY PHOTO Anything that is captured will be held agers captured a female cougar and The Wyoming Game and Fish Department officials received hundreds of pages in a bear trap in our shop so as not to her three kittens after they repeatedly of comments from people asking them to return a female cougar and her break up the group. However, we can- killed deer near private homes near kittens to the Cache Creek drainage. Wildlife managers captured the cats and not hold them for long, so we will have relocated them south of Lander after they killed deer on private property.