Leaf Traits of Drought Tolerance for 37 Shrub Species Originating from a Moisture Gradient
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Banksia Serrata ‘Superman’
NNativeNativeative PPlantsPlantslants fforforor NNewNewew SSouthSouthouth WWalesWalesales JJournalournal ooff tthehe AAustralianustralian PPlantslants SocietySociety NNSWSWNative LtdLtd VolVPlantsol 4848 April NoNo 2013 2 AprilA —pr Pageil 201320 13 Native Plants for NSW Published quarterly in January, April, July and Contents October by the Australian Plants Society NSW Introduction ...................................... 3 Ltd ACN 002 680 408 Quarterly Meeting at Menai Group .. 4 Editor: David Crawford Establishing a Native Grass Lawn ...6 Proof Reading: Rhonda Daniels Jan Douglas Merimbula Star-Hair ....................... 9 Layout: Lachlan McLaine DIY Insect Hotels ...........................10 The Journal is a forum for the exchange President’s report .......................... 12 of views of members and others and their Coates Wildlife Tours .....................13 experiences of propagating, conserving and gardening with Australian plants. Our Bird-friendly Garden ............... 14 Contributions are warmly welcomed. They Vale Isobel Hoegh-Guldberg ......... 16 may be typed or handwritten and accompanied by photographs and drawings. If handwritten, Acacia pubifolia (TTSG) ................ 17 please print botanical names and names of APS NSW Annual .......................... 18 people. General Meeting Photographs may be submitted as either high Vale Noel Gane ............................. 20 resolution digital ¿ les, such as jpg, or prints. New members ............................... 21 APS NSW Of¿ ce Mail: PO Box 5026 2013 Annual -

Sunvalley Plants Nursery
SUNVALLEY PLANTS NURSERY 7cm Tubes Bauera 'Candy Stripe' Beaufortia sparsa Chorizema cordatum Chrysocephalum apiculatum Correa decumbens Correa 'Fat Fred' Correa 'Little Cate' Correa 'Pink Mist' Correa reflexa (green) Dampiera diversifolia Dampiera trigona Eremophila 'Amber Carpet' Eremophila calorhabdos Eremophila 'Kalbarri Carpet' Eremophila 'Lime Gold' Eremophila maculata (apricot) Eremophila maculata (purple) Eremophila 'Mingenew Gold' Eremophila oppositifolia (pink) Eremophila 'Passion Pink' Eremophila 'Red Desert' Eremophila 'Wild Berry' Eutaxia obovata Grevillea 'Baby Face' Grevillea 'Emma Charlotte' Grevillea obtusifolia Grevillea x gaudichaudii Grevillea x semperflorens Lechenaultia 'Lola Pink' Leptospermum 'Pink Cascade' Myoporum parvifolium (pink) Myoporum parvifolium (purple) Pimelea ferruginea (pink) Pimelea ferruginea alba Pimelea nivea Tetratheca thymifolia Zieria 'Carpet Star' 14cm pots Anigozanthus ‘Red Volcano’ Banksia speciosa Blandfordia grandiflora Callistemon ‘Red Alert’ Callistemon ‘Scarlet Flame’ Correa ‘Fat Fred’ Correa ‘Pink Frost’ Correa alba Correa pulchella Eucalyptus macrandra Eucalyptus sideroxylon rosea Eucalyptus torquata Grevillea ‘Alpine Beauty’ Grevillea ‘Fancy Nancy’ Grevillea ‘Gold Rush’ Grevillea ‘Lady O’ Grevillea ‘Robyn Gordon’ Grevillea ‘Shades of Green’ Grevillea ‘Winter Wonder’ Grevillea juniperina Grevillea x gaudichaudii Guichenotia macrantha Pimelea nivea 20cm pots Banksia ‘Birthday Candles’ Banksia ‘Black Magic’ Conostylus aculeata Grevillea ‘Caffachino’ Grevillea x gaudichaudii Grevillea nudiflora Grevillea ‘Royal Mantle’ Grevillea ‘Rowdy’ Hardenbergia ‘Happy Wanderer’ Grafted Short Standards Grevillea ‘Droopy Drawers’ Grevillea 'Royal Mantle' Grevillea ‘Sunkissed Waters’ Low Grafts Acacia baileyana (prostrate) Grevillea ‘Bundle O Joy’ Grevillea ‘Bush Lemons’ Grevillea ‘Crowning Glory’ Grevillea ‘Ivory Whip’ Grevillea flexuosa Grafted Tall Standards Callistemon ‘Firebrand’ Grevillea ‘Fanfair’ Grevillea thyrsoides Grafted Eucalyptus Eucalyptus dwarf orange, red & pink.. -
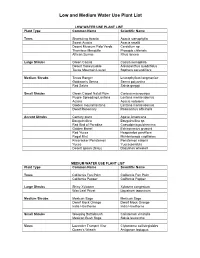
Low and Medium Water Use Plant List
Low and Medium Water Use Plant List LOW WATER USE PLANT LIST Plant Type Common Name Scientific Name Trees Shoestring Acacia Acacia stenophylla Sweet Acacia Acacia smallii Desert Museum Palo Verde Cercidium sp. Thornless Mesquite Prosopis chilensis African Sumac Rhus lancea Large Shrubs Green Cassia Cassia nemophila Desert Honeysuckle Anisacanthus quadrifidus Texas Mountain Laurel Sophora secundiflora Medium Shrubs Texas Ranger Leucophyllum langmaniae Goldman’s Senna Senna polyantha Red Salvia Salvia greggii Small Shrubs Green Carpet Natal Plum Carissa macrocarpa Purple Spreading Lantana Lantana montevidensis Acacia Acacia redolens Golden mound lantana Lantana montevidensis Dwarf Rosemary Rosmarinus officinalis Accent Shrubs Century plant Agave Americana Bougainvillea Bougainvillea sp. Red Bird of Paradise Caesalpinia pulcherrima Golden Barrel Echinocactus grusonii Red Yucca Hesperaloe parviflora Regal Mist Muhlenbergia capillaries Firecracker Penstemon Penstemon eatonii Yucca Yucca pendula Desert spoon (Grey) Dasylirion wheeleri MEDIUM WATER USE PLANT LIST Plant Type Common Name Scientific Name Trees California Fan Palm California Fan Palm California Pepper California Pepper Large Shrubs Shiny Xylosma Xylosma congestum Wax Leaf Privet Ligustrum japonicum Medium Shrubs Mexican Sage Mexican Sage Dwarf Mock Orange Dwarf Mock Orange India Hawthorne India Hawthorne Small Shrubs Weeping Bottlebrush Calistemon viminalis Mexican Bush Sage Salvia leucantha Vines Lavender Trumpet Vine Clytostoma callistegioides Queen’s Wreath Antigonon leptopus . -

PUBLISHER S Candolle Herbarium
Guide ERBARIUM H Candolle Herbarium Pamela Burns-Balogh ANDOLLE C Jardin Botanique, Geneva AIDC PUBLISHERP U R L 1 5H E R S S BRILLB RI LL Candolle Herbarium Jardin Botanique, Geneva Pamela Burns-Balogh Guide to the microform collection IDC number 800/2 M IDC1993 Compiler's Note The microfiche address, e.g. 120/13, refers to the fiche number and secondly to the individual photograph on each fiche arranged from left to right and from the top to the bottom row. Pamela Burns-Balogh Publisher's Note The microfiche publication of the Candolle Herbarium serves a dual purpose: the unique original plants are preserved for the future, and copies can be made available easily and cheaply for distribution to scholars and scientific institutes all over the world. The complete collection is available on 2842 microfiche (positive silver halide). The order number is 800/2. For prices of the complete collection or individual parts, please write to IDC Microform Publishers, P.O. Box 11205, 2301 EE Leiden, The Netherlands. THE DECANDOLLEPRODROMI HERBARIUM ALPHABETICAL INDEX Taxon Fiche Taxon Fiche Number Number -A- Acacia floribunda 421/2-3 Acacia glauca 424/14-15 Abatia sp. 213/18 Acacia guadalupensis 423/23 Abelia triflora 679/4 Acacia guianensis 422/5 Ablania guianensis 218/5 Acacia guilandinae 424/4 Abronia arenaria 2215/6-7 Acacia gummifera 421/15 Abroniamellifera 2215/5 Acacia haematomma 421/23 Abronia umbellata 221.5/3-4 Acacia haematoxylon 423/11 Abrotanella emarginata 1035/2 Acaciahastulata 418/5 Abrus precatorius 403/14 Acacia hebeclada 423/2-3 Acacia abietina 420/16 Acacia heterophylla 419/17-19 Acacia acanthocarpa 423/16-17 Acaciahispidissima 421/22 Acacia alata 418/3 Acacia hispidula 419/2 Acacia albida 422/17 Acacia horrida 422/18-20 Acacia amara 425/11 Acacia in....? 423/24 Acacia amoena 419/20 Acacia intertexta 421/9 Acacia anceps 419/5 Acacia julibross. -

Cunninghamia Date of Publication: February 2020 a Journal of Plant Ecology for Eastern Australia
Cunninghamia Date of Publication: February 2020 A journal of plant ecology for eastern Australia ISSN 0727- 9620 (print) • ISSN 2200 - 405X (Online) The Australian paintings of Marianne North, 1880–1881: landscapes ‘doomed shortly to disappear’ John Leslie Dowe Australian Tropical Herbarium, James Cook University, Smithfield, Qld 4878 AUSTRALIA. [email protected] Abstract: The 80 paintings of Australian flora, fauna and landscapes by English artist Marianne North (1830-1890), completed during her travels in 1880–1881, provide a record of the Australian environment rarely presented by artists at that time. In the words of her mentor Sir Joseph Dalton Hooker, director of Kew Gardens, North’s objective was to capture landscapes that were ‘doomed shortly to disappear before the axe and the forest fires, the plough and the flock, or the ever advancing settler or colonist’. In addition to her paintings, North wrote books recollecting her travels, in which she presented her observations and explained the relevance of her paintings, within the principles of a ‘Darwinian vision,’ and inevitable and rapid environmental change. By examining her paintings and writings together, North’s works provide a documented narrative of the state of the Australian environment in the late nineteenth- century, filtered through the themes of personal botanical discovery, colonial expansion and British imperialism. Cunninghamia (2020) 20: 001–033 doi: 10.7751/cunninghamia.2020.20.001 Cunninghamia: a journal of plant ecology for eastern Australia © 2020 Royal Botanic Gardens and Domain Trust www.rbgsyd.nsw.gov.au/science/Scientific_publications/cunninghamia 2 Cunninghamia 20: 2020 John Dowe, Australian paintings of Marianne North, 1880–1881 Introduction The Marianne North Gallery in the Royal Botanic Gardens Kew houses 832 oil paintings which Marianne North (b. -

Acacia Fimbriata Dwarf Crimson Blush 8 Eye on It During the Conference, Please Let Me Know
Australian Native Plants Society (Australia) Inc. ACACIA STUDY GROUP NEWSLETTER Group Leader and Newsletter Editor Seed Bank Curator Bill Aitchison Victoria Tanner 13 Conos Court, Donvale, Vic 3111 Phone (03) 98723583 Email: [email protected] No. 129 June 2015 ISSN 1035-4638 Contents Page From The Leader Dear Members From the Leader 1 It is now only a few months until the ANPSA Biennial Welcome 2 Conference being held in Canberra from 15-20 November. From Members and Readers 2 This is a great opportunity to catch up with some other Some Notes From Yallaroo 3 members of our Study Group, and of course to take part in Wattles With Minni Ritchi Bark 5 the great program put together by the organisers. Introduction of Australian Acacias Information relating to the Conference and details regarding to South America 6 registration are available on the Conference website Max’s Interesting Wattles 7 http://anpsa.org.au/conference2015. Our Study Group will An Acacia dealbata question from have a display at the Conference. If any Study Group Sweden 7 member who will be at the Conference could help with the Pre-treatment of Acacia Seeds 8 display, either in setting it up, or just in helping to keep an Acacia fimbriata dwarf Crimson Blush 8 eye on it during the Conference, please let me know. Books 9 Seed Bank 9 I am sure that many of our members will be aware of the Study Group Membership 10 Wattle Day Association, and the great work that it does in promoting National Wattle Day each year on 1 September. -

Newsletter No
Newsletter No. 167 June 2016 Price: $5.00 AUSTRALASIAN SYSTEMATIC BOTANY SOCIETY INCORPORATED Council President Vice President Darren Crayn Daniel Murphy Australian Tropical Herbarium (ATH) Royal Botanic Gardens Victoria James Cook University, Cairns Campus Birdwood Avenue PO Box 6811, Cairns Qld 4870 Melbourne, Vic. 3004 Australia Australia Tel: (+61)/(0)7 4232 1859 Tel: (+61)/(0) 3 9252 2377 Email: [email protected] Email: [email protected] Secretary Treasurer Leon Perrie John Clarkson Museum of New Zealand Te Papa Tongarewa Queensland Parks and Wildlife Service PO Box 467, Wellington 6011 PO Box 975, Atherton Qld 4883 New Zealand Australia Tel: (+64)/(0) 4 381 7261 Tel: (+61)/(0) 7 4091 8170 Email: [email protected] Mobile: (+61)/(0) 437 732 487 Councillor Email: [email protected] Jennifer Tate Councillor Institute of Fundamental Sciences Mike Bayly Massey University School of Botany Private Bag 11222, Palmerston North 4442 University of Melbourne, Vic. 3010 New Zealand Australia Tel: (+64)/(0) 6 356- 099 ext. 84718 Tel: (+61)/(0) 3 8344 5055 Email: [email protected] Email: [email protected] Other constitutional bodies Hansjörg Eichler Research Committee Affiliate Society David Glenny Papua New Guinea Botanical Society Sarah Matthews Heidi Meudt Advisory Standing Committees Joanne Birch Financial Katharina Schulte Patrick Brownsey Murray Henwood David Cantrill Chair: Dan Murphy, Vice President Bob Hill Grant application closing dates Ad hoc adviser to Committee: Bruce Evans Hansjörg Eichler Research -

Downloading Or Purchasing Online At
On-farm Evaluation of Grafted Wildflowers for Commercial Cut Flower Production OCTOBER 2012 RIRDC Publication No. 11/149 On-farm Evaluation of Grafted Wildflowers for Commercial Cut Flower Production by Jonathan Lidbetter October 2012 RIRDC Publication No. 11/149 RIRDC Project No. PRJ-000509 © 2012 Rural Industries Research and Development Corporation. All rights reserved. ISBN 978-1-74254-328-4 ISSN 1440-6845 On-farm Evaluation of Grafted Wildflowers for Commercial Cut Flower Production Publication No. 11/149 Project No. PRJ-000509 The information contained in this publication is intended for general use to assist public knowledge and discussion and to help improve the development of sustainable regions. You must not rely on any information contained in this publication without taking specialist advice relevant to your particular circumstances. While reasonable care has been taken in preparing this publication to ensure that information is true and correct, the Commonwealth of Australia gives no assurance as to the accuracy of any information in this publication. The Commonwealth of Australia, the Rural Industries Research and Development Corporation (RIRDC), the authors or contributors expressly disclaim, to the maximum extent permitted by law, all responsibility and liability to any person, arising directly or indirectly from any act or omission, or for any consequences of any such act or omission, made in reliance on the contents of this publication, whether or not caused by any negligence on the part of the Commonwealth of Australia, RIRDC, the authors or contributors. The Commonwealth of Australia does not necessarily endorse the views in this publication. This publication is copyright. -

Ne Wsletter No . 92
AssociationAustralian of NativeSocieties Plants for Growing Society (Australia)Australian IncPlants Ref No. ISSN 0725-8755 Newsletter No. 92 – August 2012 GSG Vic Programme 2012 GSG SE Qld Programme 2012 Leader: Neil Marriott Morning tea at 9.30am, meetings commence at 693 Panrock Reservoir Rd, Stawell, Vic. 3380 10.00am. For more information contact Bryson Phone: 03 5356 2404 or 0458 177 989 Easton on (07) 3121 4480 or 0402242180. Email: [email protected] Sunday, 26 August Contact Neil for queries about program for the year. This meeting has been cancelled as many members Any members who would like to visit the official have another function to attend over the weekend. collection, obtain cutting material or seed, assist in its maintenance, and stay in our cottage for a few days The October 2012 meeting – has been are invited to contact Neil. After the massive rains at replaced by a joint excursion through SEQ & the end of 2010 and the start of 2011 the conditions northern NSW commencing on Wednesday, 7 are perfect for large scale replanting of the collection. November 2012. GSG members planning to attend Offers of assistance would be most welcome. are asked to contact Jan Glazebrook & Dennis Cox Newsletter No. 92 No. Newsletter on Ph (07) 5546 8590 for full details closer to this Friday, 29 September to Monday, 1 October event. See also page 3 for more details. SUBJECT: Spring Grevillea Crawl Sunday, 25 November FRI ARVO: Meet at Neil and Wendy Marriott’s Panrock VENUE: Home of Robyn Wieck Ridge, 693 Panrock Reservoir Rd, Stawell Lot 4 Ajuga Court, Brookvale Park Oakey for welcome and wander around the HONE (07) 4691 2940 gardens. -

Verticordia Study Group
S.G.1.P. QLD. REGION - LIBRARY - ASSOCIATION OF SOCIETIES FOR GROWING AUSTRALIAN PLANTS VERTICORDIA STUDY GROUP NEWSLETER NO 44 -- July 2005. MEMBERSHIP I am very pleased to welcome the following new members to our Study Group :- Bob Carroll, 23 Perigee Close, Doonside, NSW 2767 Bob comments:- "I know little about Verticordias except that they are mostly reputed to be acultto grow in my area; (Sydney"~outer western suburbs). I have had some success in the past growhg them in containers, but even then they did not live long, probably due to lack of skill and knowledge on my part. but I wish to learn more and would appreciate your advice" Ian Budge, RSM 6, Biddle Road, Dunsborough, W.A. 6281 Ian lives on a 100 acre property, 250 Km south of Perth and 6 Km fiom the sea.. He works as a Coordinator for Parks and Gardens in the Busselton Shire and says " My training in horticulture some 26 years ago still fuels the passion I have for W.A. Native Plants but my work rarely involves verticordia. My interest in verticordia can be blamed on Elizabeth A. George's book 'Verticordia, the Turner of Hearts' ;.an excellent layman's guide to this genus.-------I had the pleasure of meeting her in person at the Wildflower Society Landsdale Nursery where I was able to pick up a few rare and endangered species and try them out on our property. ,4bout 14 species have been planted in the last 12 months on our grey sand overlaying gravelly soil.-----They are doing well with occasional light watering during hot weather and protected by planter bags fiom clumsy roos and damaging rabbits. -

TREES Botanical Common Acacia Aneura Mulga Acacia Berlandieri
TREES Botanical Common Acacia aneura Mulga Acacia berlandieri Guajillo Acacia craspedocarpa Leatherleaf Acacia Acacia farnesiana Sweet Acacia Acacia rigidula Blackbrush Acacia Acacia salicina Willow Acacia Acacia saligna Blue Leaf Wattle Acacia stenophylla Shoestring Acacia Acacia willardiana Palo Blanco Albizia julibrissin Silk tree, Mimosa Tree Arecastrum romanzoffianum Queen Palm Bauhinia blakeana Hong Kong Orchid Tree Bauhinia lunarioides White Orchid Tree Bauhinia Purpurea Purple Orchid Tree Bauhinia variegata Purple Orchid Tree Brachychiton populneus Bottle Tree Brahea armata Mexican Blue Palm Brahea edulis Guadalupe Island Palm Butia Capitata Pindo Palm Caesalpinia cacalaco Cascalote Callistemon viminalis Bottle Brush Tree Ceratonia siliqua Carob Tree Chamaerops humilis Mediterranean Fan Palm Chilopsis linearis Desert Willow Chitalpa X tashkentenis Chitalpa Chorisia speciosa Silk floss Tree, Kapok Cupressus arizonica Arizona Cypress Cupressus Sempervirens Italian Cypress Dalbergia sissoo Indian Rosewood Dalea spinosa Desert Smoke Tree Eriobotrya japonica Loquat, Japanese Plum Eucalyptus cinerea Silver-Dollar Tree Eucalyptus krusaena Kruses Eucalyptus Eucalyptus microtheca Coolibah Tree Eucalyptus papuana Ghost Gum Eucalyptus spathulata Swamp Mallee Eysenhardtia orthocarpa Kidneywood Fraxinus uhdei Evergreen Ash Geijera parviflora Australian Willow Jacaranda mimosifolia Jacaranda Koelreuteria bipinnata Chinese Flame Tree Lagerstroemia indica Crape Myrtle Lysiloma watsonii var. thornberi Feather Tree Melaleuca quinquenervia Cajeput -

Leather Leaf Acacia, Wax Leaf Acacia Flower Color: Bloom Season: Spring to Summer Water Use - 1: Indicates Very Low Water Use
1 BOTANICAL NAME: Family: Fabaceae Acacia craspedocarpa COMMON NAME: Leather Leaf Acacia, Wax Leaf Acacia Flower Color: Bloom season: Spring to Summer Water Use - 1: indicates very low water use. After establishment water every 3 - 6 weeks during the growing season. Growing season: Winter Apply water September through March; less frequently in off season. Picture: Rich Benda During hot summer months this plant may need to be watered more often. Characteristics: Landscape Use: Width: 12’ Height: 12’ (50% to 75% mature size) Toxic: No An evergreen that is used best for hedge or Allergenicity: Weakly screen. Invasive: No Spreads in cultivated areas: No Cold Hardiness: 0F 18 Growing Conditions: Elevation: 1,000’ to 5,000’ It likes full sun, takes heat Origin: Australia Soil tolerant with good drainage. Information Sources Low Water Use/Drought Tolerant Plant List - Official Regulatory List For: - Arizona Department of Water Resources - Tucson Active Management Area Plants for Dry Climates: Duffield, Mary Rose and Jones, Warren - Revised Edition © 2001 A Field Guide to the Plants of Arizona: Epple, Anne Orth - First Edition / Tenth Printing © 1995 Native Plants for Southwestern Landscapes: Mielke, Judy - © 1993 Sixth paperback printing, 2005 2 BOTANICAL NAME: Family: Asteraceae Ambrosia deltoidea (Franseria) COMMON NAME: Triangle-leaf Bursage Flower Color: Inconspicuous Bloom season: Mid-Winter to Mid Spring Water Use - 1: indicates very low water use. After establishment water every 3 - 6 weeks during the growing season. Growing season: Winter Apply water September through March; less frequently in off season. Picture: Rich Benda During hot summer months this plant may need to be watered more often.