Nymphalidae) in Southern Arizona
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Phylogenetic Relationships and Historical Biogeography of Tribes and Genera in the Subfamily Nymphalinae (Lepidoptera: Nymphalidae)
Blackwell Science, LtdOxford, UKBIJBiological Journal of the Linnean Society 0024-4066The Linnean Society of London, 2005? 2005 862 227251 Original Article PHYLOGENY OF NYMPHALINAE N. WAHLBERG ET AL Biological Journal of the Linnean Society, 2005, 86, 227–251. With 5 figures . Phylogenetic relationships and historical biogeography of tribes and genera in the subfamily Nymphalinae (Lepidoptera: Nymphalidae) NIKLAS WAHLBERG1*, ANDREW V. Z. BROWER2 and SÖREN NYLIN1 1Department of Zoology, Stockholm University, S-106 91 Stockholm, Sweden 2Department of Zoology, Oregon State University, Corvallis, Oregon 97331–2907, USA Received 10 January 2004; accepted for publication 12 November 2004 We infer for the first time the phylogenetic relationships of genera and tribes in the ecologically and evolutionarily well-studied subfamily Nymphalinae using DNA sequence data from three genes: 1450 bp of cytochrome oxidase subunit I (COI) (in the mitochondrial genome), 1077 bp of elongation factor 1-alpha (EF1-a) and 400–403 bp of wing- less (both in the nuclear genome). We explore the influence of each gene region on the support given to each node of the most parsimonious tree derived from a combined analysis of all three genes using Partitioned Bremer Support. We also explore the influence of assuming equal weights for all characters in the combined analysis by investigating the stability of clades to different transition/transversion weighting schemes. We find many strongly supported and stable clades in the Nymphalinae. We are also able to identify ‘rogue’ -

Origins of Six Species of Butterflies Migrating Through Northeastern
diversity Article Origins of Six Species of Butterflies Migrating through Northeastern Mexico: New Insights from Stable Isotope (δ2H) Analyses and a Call for Documenting Butterfly Migrations Keith A. Hobson 1,2,*, Jackson W. Kusack 2 and Blanca X. Mora-Alvarez 2 1 Environment and Climate Change Canada, 11 Innovation Blvd., Saskatoon, SK S7N 0H3, Canada 2 Department of Biology, University of Western Ontario, Ontario, ON N6A 5B7, Canada; [email protected] (J.W.K.); [email protected] (B.X.M.-A.) * Correspondence: [email protected] Abstract: Determining migratory connectivity within and among diverse taxa is crucial to their conservation. Insect migrations involve millions of individuals and are often spectacular. However, in general, virtually nothing is known about their structure. With anthropogenically induced global change, we risk losing most of these migrations before they are even described. We used stable hydrogen isotope (δ2H) measurements of wings of seven species of butterflies (Libytheana carinenta, Danaus gilippus, Phoebis sennae, Asterocampa leilia, Euptoieta claudia, Euptoieta hegesia, and Zerene cesonia) salvaged as roadkill when migrating in fall through a narrow bottleneck in northeast Mexico. These data were used to depict the probabilistic origins in North America of six species, excluding the largely local E. hegesia. We determined evidence for long-distance migration in four species (L. carinenta, E. claudia, D. glippus, Z. cesonia) and present evidence for panmixia (Z. cesonia), chain (Libytheana Citation: Hobson, K.A.; Kusack, J.W.; Mora-Alvarez, B.X. Origins of Six carinenta), and leapfrog (Danaus gilippus) migrations in three species. Our investigation underlines Species of Butterflies Migrating the utility of the stable isotope approach to quickly establish migratory origins and connectivity in through Northeastern Mexico: New butterflies and other insect taxa, especially if they can be sampled at migratory bottlenecks. -

Arthropods of Elm Fork Preserve
Arthropods of Elm Fork Preserve Arthropods are characterized by having jointed limbs and exoskeletons. They include a diverse assortment of creatures: Insects, spiders, crustaceans (crayfish, crabs, pill bugs), centipedes and millipedes among others. Column Headings Scientific Name: The phenomenal diversity of arthropods, creates numerous difficulties in the determination of species. Positive identification is often achieved only by specialists using obscure monographs to ‘key out’ a species by examining microscopic differences in anatomy. For our purposes in this survey of the fauna, classification at a lower level of resolution still yields valuable information. For instance, knowing that ant lions belong to the Family, Myrmeleontidae, allows us to quickly look them up on the Internet and be confident we are not being fooled by a common name that may also apply to some other, unrelated something. With the Family name firmly in hand, we may explore the natural history of ant lions without needing to know exactly which species we are viewing. In some instances identification is only readily available at an even higher ranking such as Class. Millipedes are in the Class Diplopoda. There are many Orders (O) of millipedes and they are not easily differentiated so this entry is best left at the rank of Class. A great deal of taxonomic reorganization has been occurring lately with advances in DNA analysis pointing out underlying connections and differences that were previously unrealized. For this reason, all other rankings aside from Family, Genus and Species have been omitted from the interior of the tables since many of these ranks are in a state of flux. -
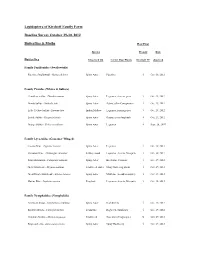
Butterflies and Moths List
Lepidoptera of Kirchoff Family Farm Baseline Survey October 29-30, 2012 Butterflies & Moths Host Plant Species Present Date Butterflies Observed On Larval Host Plants Kirchoff FF observed Family Papilionidae (Swallowtails) Pipevine Swallowtail - Battus philenor Spiny Aster Pipevine √ Oct. 30, 2012 Family Pieridae (Whites & Sulfurs) Cloudless Sulfur - Phoebis sennae Spiny Aster Legumes, clovers, peas √ Oct. 29, 2012 Dainty Sulfur - Nathalis iole Spiny Aster Asters, other Com;posites √ Oct. 29, 2012 Little Yellow Sulfur - Eurema lisa Indian Mallow Legumes, partridge pea √ Oct. 29, 2012 Lyside Sulfur - Kirgonia lyside Spiny Aster Guayacan or Soapbush √ Oct. 29, 2012 Orange Sulfur - Colias eurytheme Spiny Aster Legumes √ Sept. 26, 2007 Family Lycaenidae (Gossamer Winged) Cassius Blue - Leptotes cassius Spiny Aster Legumes √ Oct. 30, 2012 Ceraunus Blue - Hemiargus ceraunus Kidney wood Legumes- Acacia, Mesquite √ Oct. 30, 2012 Fatal Metalmark - Calephelis nemesis Spiny Aster Baccharis, Clematis √ Oct. 29, 2012 Gray Hairstreak - Strymon melinus Frostweed, Aster Many flowering plants √ Oct. 29, 2012 Great Purple Hairstreak - Atlides halesus Spiny Aster Mistletoe (in oak/mesquite) √ Oct. 29, 2012 Marine Blue - Leptotes marina Frogfruit Legumes- Acacia, Mesquite √ Oct. 30, 2012 Family Nymphalidae (Nymphalids) American Snout - Libytheana carinenta Spiny Aster Hackberries √ Oct. 29, 2012 Bordered Patch - Chlosyne lacinia Zexmenia Ragweed, Sunflower √ Oct. 29, 2012 Common Mestra - Mestra amymone Frostweed Noseburn (Tragia spp.) X Oct. 29, 2012 Empress Leila - Asterocampa leilia Spiny Aster Spiny Hackberry √ Oct. 29, 2012 Gulf Fritillary - Agraulis vanillae Spiny Aster Passion Vine X Oct. 29, 2012 Hackberry Emperor - Asterocampa celtis spiny Hackberry Hackberries √ Oct. 29, 2012 Painted Lady - Vanessa cardui Pink Smartweed Mallows & Thistles √ Oct. 29, 2012 Pearl Crescent - Phycoides tharos Spiny Aster Asters √ Oct. -

The First Record of Aglais Milberti (Godart, 1819) from the Territory of Russia 101 Atalanta 42 (1-4): 101, Würzburg (2011), ISSN 0171-0079
ZOBODAT - www.zobodat.at Zoologisch-Botanische Datenbank/Zoological-Botanical Database Digitale Literatur/Digital Literature Zeitschrift/Journal: Atalanta Jahr/Year: 2011 Band/Volume: 42 Autor(en)/Author(s): Churkin Sergei V., Zamolodchikov Dmitry Artikel/Article: The first record of Aglais milberti (Godart, 1819) from the territory of Russia 101 Atalanta 42 (1-4): 101, Würzburg (2011), ISSN 0171-0079 The first record ofAglais milberti (GODART , 1819) from the territory of Russia (Lepidoptera, Nymphalidae) by SERGEI CHURKIN & DMITRY ZAMOLO dc HIKOV received 20.V.2011 Summary: Aglais milberti (GO D ART , 1819) was found in the territory of Russia (East Churkotka) for the first time. The subspecies status needs further clarification. Резюме: Aglais milberti (GO D ART , 1819) впервые отмечена для территории России (Вост. Чукотка). Подвидовой статус нуждается в дальнейшем уточнении. A fresh † of Aglais milberti (GO D ART , 1819) (the figures are enlarged) was collected by .D ZAMOLO dc HIKOV at Lavrentia Bay (Russia, East Churkotka, 20 km W Lavrentia Bay , “Tri brata” hill, 400 m.a.s.l. , 27.VII.2004). It is deposited in the private collection of the second author. Worth to note several facts: - there are no Aglais-records from Chukotka; - this is the first record of this species from the territory of the Russian Federation; - the butterfly was found in the place where the second author collected many times; the butterflies were not found before 2004 neither after 2004. The habitat is typical for Chukotka: alpine meadows in closed valleys, protected from the wind by the surrounding mountain slopes; the meadows are situated among the tundras with Betula exilis. -

Butterflies in the Verde Valley
Butterflies in the Verde Valley 1 2 17 1. Mormon Fritillary, Speyeria mormonia 2. Empress Leilia, Asterocampa leilia 3. Fiery Skipper, Hylephila phyleus 4. Greenish-Blue Lycaenid, Plebejus saepiolus 16 4 18 Female on top, male below 3 5. Pipevine Swallowtail, Battus philenor 6. American Snout Butterfly, Libytheana carinenta 7. Cloudless Sulpher, Phoebis sennae 14 15 Female with patterned wing, male solid 5 8. Two-tailed Swallowtail, Papilio multicaudata 9. Variegated Fritillary, Euptoieta claudia 13 12 10. Hoary Comma, Polygonia gracilis 6 11. California Sister, Adelpha bredowii Eurema nicippe 8 12. Sleepy Orange Sulphur, 11 Two males 10 13. Alfalfa Sulfpur, Colias euretheme Two males, one female (pale) 14. Painted Lady, Vanessa cardui 15. Pine White, Neophasia menapia 9 7 16. Viceroy, Limenitus archippus 17. Queen, Danaus gilippus 18. Black Swallowtail, Papilio polyxenes Butterflies Butterflies are an amazing group of insects, and their delicate structure, flight, and colors, brighten our day. The butterflies in this display represent 6 related families of Lepidoptera (scaled wings) which we know as butterflies. All of these occur in northern Arizona. The best place to look for butterflies is often in moist creek beds and in areas where flowers are blooming. The mouth is composed of two long flexible straws that are connected. When they feed/drink they unroll the mouth parts (called a proboscis) and suck in nectar or other liquids. Although the shapes and colors differs, their basic structure is similar. All have four wings, six legs, an abdomen, thorax, head, and antenna. Butterflies have complete metamorphosis, with four separate stages: (1)egg, (2)larva, (3)chrysalis, and (4)adult. -

Phylogenetic Relatedness of Erebia Medusa and E. Epipsodea (Lepidoptera: Nymphalidae) Confirmed
Eur. J. Entomol. 110(2): 379–382, 2013 http://www.eje.cz/pdfs/110/2/379 ISSN 1210-5759 (print), 1802-8829 (online) Phylogenetic relatedness of Erebia medusa and E. epipsodea (Lepidoptera: Nymphalidae) confirmed 1 2, 3 4 MARTINA ŠEMELÁKOVÁ , PETER PRISTAŠ and ĽUBOMÍR PANIGAJ 1 Institute of Biology and Ecology, Department of Cellular Biology, Faculty of Science, Pavol Jozef Šafárik University in Košice, Moyzesova 11, 041 54 Košice, Slovakia; e-mail: [email protected] 2 Institute of Animal Physiology, Slovak Academy of Science, Soltesovej 4–6, 040 01 Košice, Slovakia 3 Department of Biology and Ecology, Faculty of Natural Sciences, Matej Bel University, Tajovskeho 40, 841 04 Banská Bystrica, Slovakia 4 Institute of Biology and Ecology, Department of Zoology, Faculty of Science, Pavol Jozef Šafárik University in Košice, Moyzesova 11, 041 54 Košice, Slovakia Key words. Lepidoptera, Nymphalidae, Erebia medusa, E. epipsodea, mtDNA, COI, ND1 Abstract. The extensive genus Erebia is divided into several groups of species according to phylogenetic relatedness. The species Erebia medusa was assigned to the medusa group and E. epipsodea to the alberganus group. A detailed study of the morphology of their copulatory organs indicated that these species are closely related and based on this E. epipsodea was transferred to the medusa group. Phylogenetic analyses of the gene sequences of mitochondrial cytochrome C oxidase subunit I (COI) and mitochondrial NADH dehydrogenase subunit 1 (ND1) confirm that E. medusa and E. epipsodea are closely related. A possible scenario is that the North American species, E. episodea, evolved after exclusion/isolation from E. medusa, whose current centre of distribution is in Europe. -

New Distribution Record of Indian Tortoiseshell Aglais Caschmirensis Aesis (Fruhstorfer, 1912) (Lepidoptera: Nymphalidae) from the State of Assam, India
www.biotaxa.org/rce. ISSN 0718-8994 (online) Revista Chilena de Entomología (2021) 47 (2): 253-258. Scientific Note New distribution record of Indian Tortoiseshell Aglais caschmirensis aesis (Fruhstorfer, 1912) (Lepidoptera: Nymphalidae) from the state of Assam, India Nuevo registro de distribución de Aglais caschmirensis aesis (Fruhstorfer, 1912) (Lepidoptera: Nymphalidae) del estado de Assam, India Debaprasad Sengupta1 ,2 , Subhasish Arandhara2 and Puja Deb2 1Bansbari Pathar, Dibrugarh- 786001, Assam, India. 2Department of Zoology & Wildlife Biology, A.V.C. College (Autonomous), Mannampandal- 609305, Mayiladuthurai, Tamil Nadu, India. debaprasad. [email protected] ZooBank: urn:lsid:zoobank.org:pub:5FBE5E7E-71A8-4309-B21C-1E9B4E811D35 https://doi.org/10.35249/rche.47.2.21.12 Abstract. Opportunistic sightings of Aglais caschmirensis aesis were recorded for the first time (on two different occasions) from Dibrugarh city in the state of Assam with new elevation records (109 and 113 m) from the north-eastern region of India. Key words: Butterfly; Dibrugarh; Jokai Reserve Forest; Northeast India; Tea Estate. Resumen. Los avistamientos de Aglais caschmirensis aesis se realizaron por primera vez (en dos ocasiones diferentes) en la ciudad de Dibrugarh en el estado de Assam con nuevos registros de elevación (109 y 113 m) en la región noreste de la India. Palabras clave: Mariposa; Dibrugarh; Reserva Forestal de Jokai; Tea Estate; noreste de India. North-eastern region of India comprises of Arunachal Pradesh, Assam, Manipur, Meghalaya, Mizoram, Nagaland, Sikkim, Tripura, and Northern parts of West Bengal (Choudhury 2004) and is one of the richest biodiversity zones of India which harbors around 64% of the total butterfly species reported from India (Joshi & Dhyani 2014). -

BUTTERFLIES in Thewest Indies of the Caribbean
PO Box 9021, Wilmington, DE 19809, USA E-mail: [email protected]@focusonnature.com Phone: Toll-free in USA 1-888-721-3555 oror 302/529-1876302/529-1876 BUTTERFLIES and MOTHS in the West Indies of the Caribbean in Antigua and Barbuda the Bahamas Barbados the Cayman Islands Cuba Dominica the Dominican Republic Guadeloupe Jamaica Montserrat Puerto Rico Saint Lucia Saint Vincent the Virgin Islands and the ABC islands of Aruba, Bonaire, and Curacao Butterflies in the Caribbean exclusively in Trinidad & Tobago are not in this list. Focus On Nature Tours in the Caribbean have been in: January, February, March, April, May, July, and December. Upper right photo: a HISPANIOLAN KING, Anetia jaegeri, photographed during the FONT tour in the Dominican Republic in February 2012. The genus is nearly entirely in West Indian islands, the species is nearly restricted to Hispaniola. This list of Butterflies of the West Indies compiled by Armas Hill Among the butterfly groupings in this list, links to: Swallowtails: family PAPILIONIDAE with the genera: Battus, Papilio, Parides Whites, Yellows, Sulphurs: family PIERIDAE Mimic-whites: subfamily DISMORPHIINAE with the genus: Dismorphia Subfamily PIERINAE withwith thethe genera:genera: Ascia,Ascia, Ganyra,Ganyra, Glutophrissa,Glutophrissa, MeleteMelete Subfamily COLIADINAE with the genera: Abaeis, Anteos, Aphrissa, Eurema, Kricogonia, Nathalis, Phoebis, Pyrisitia, Zerene Gossamer Wings: family LYCAENIDAE Hairstreaks: subfamily THECLINAE with the genera: Allosmaitia, Calycopis, Chlorostrymon, Cyanophrys, -

Yukon Butterflies a Guide to Yukon Butterflies
Wildlife Viewing Yukon butterflies A guide to Yukon butterflies Where to find them Currently, about 91 species of butterflies, representing five families, are known from Yukon, but scientists expect to discover more. Finding butterflies in Yukon is easy. Just look in any natural, open area on a warm, sunny day. Two excellent butterfly viewing spots are Keno Hill and the Blackstone Uplands. Pick up Yukon’s Wildlife Viewing Guide to find these and other wildlife viewing hotspots. Visitors follow an old mining road Viewing tips to explore the alpine on top of Keno Hill. This booklet will help you view and identify some of the more common butterflies, and a few distinctive but less common species. Additional species are mentioned but not illustrated. In some cases, © Government of Yukon 2019 you will need a detailed book, such as , ISBN 978-1-55362-862-2 The Butterflies of Canada to identify the exact species that you have seen. All photos by Crispin Guppy except as follows: In the Alpine (p.ii) Some Yukon butterflies, by Ryan Agar; Cerisy’s Sphynx moth (p.2) by Sara Nielsen; Anicia such as the large swallowtails, Checkerspot (p.2) by Bruce Bennett; swallowtails (p.3) by Bruce are bright to advertise their Bennett; Freija Fritillary (p.12) by Sonja Stange; Gallium Sphinx presence to mates. Others are caterpillar (p.19) by William Kleeden (www.yukonexplorer.com); coloured in dull earth tones Butterfly hike at Keno (p.21) by Peter Long; Alpine Interpretive that allow them to hide from bird Centre (p.22) by Bruce Bennett. -

Final Lower Rio Grande Valley and Santa Ana National Wildlife
Final Lower Rio Grande Valley and Santa Ana National Wildlife Refuges Comprehensive Conservation Plan September 1997 (Reprint March 1999) U.S. Fish and Wildlife Service U.S. Department of the Interior Cover Artwork by Brian Cobble Table of Contents VISION........................................................................................................................................... 5 Executive Summary................................................................................................................... 6 1.0 Introduction and Regional Setting................................................................................. 8 1.1 LRGV Challenges............................................................................................... 8 2.0 Planning Perspectives and Considerations................................................................ 9 2.1 National Wildlife Refuge System ................................................................... 9 2.2 The Service & Ecosystem Management ...................................................... 9 2.3 Refuge Complex and Management Districts........................................... 10 2.4 Laguna Atascosa NWR -- A Partner with LRGV NWR............................ 10 2.5 Planning Perspectives.................................................................................... 10 2.6 The Issues.......................................................................................................... 11 2.7 The Need for Action........................................................................................ -

Boloria Selene) on the Ochoco National Forest
2015 Surveys for Silver-bordered Fritillary (Boloria selene) On the Ochoco National Forest Prepared by Dana Ross (Entomologist, Lepidoptera Specialist) November 2015 0 SUMMARY The silver-bordered fritillary (Boloria selene) is designated as an ISSSSP Special Status/Sensitive Species in Oregon. In June and July of 2015, surveys were performed for this regionally rare butterfly on public lands managed by the Ochoco National Forest and Bureau of Land Management where there was either historical occurrence or a likelihood of undiscovered populations of the insect. Populations were documented at 8 distinct locations from Big Summit Prairie south to Williams and Gray prairies. All observed populations were in the North Fork Crooked River drainage at sites that contained at least some seasonally wet or marshy habitats with violets (the Boloria selene larval hostplant). Three fairly sizeable populations were located – at Elliot Creek (north Big Summit Prairie), along the BLM North Fork Crooked River/lower Lookout Creek site immediately south of Big Summit Prairie, and at NW Williams Prairie. Comparatively few of the butterflies were observed elsewhere. Observed damage to fragile critical habitats, such as at Elliot Creek (off-road vehicles) and Williams Prairie (cattle grazing), must be avoided in the future to prevent habitat degradation, population decline and potential local extirpation of the insect. It is recommended that additional surveys for silver-bordered fritillary take place on National Forest and BLM lands to: 1)more completely document the species regional range and distribution; 2) to more accurately assess population/meta-population size; 3) to assess the impact to future generations from observed mid-season damage to critical habitats; 4) to verify benefits (increased abundance or local persistence) to populations from habitat protection or habitat restoration; 5) to determine butterfly response to short or long term environmental conditions such as drought.