Status Review of West Coast Steelhead from Washington
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Pacific Lamprey 2018 Regional Implementation Plan for the Lower Columbia/Willamette Regional Management Unit Lower Columbia Sub-Unit
Pacific Lamprey 2018 Regional Implementation Plan for the Lower Columbia/Willamette Regional Management Unit Lower Columbia Sub-Unit Submitted to the Conservation Team August 6th, 2018 Primary Authors Primary Editors J. Poirier, U.S. Fish and Wildlife Service This page left intentionally blank I. Status and Distribution of Pacific lamprey in the RMU A. General Description of the RMU The Lower Columbia River sub-unit within the Lower Columbia River/Willamette Regional Management Unit includes watersheds that drain into the Columbia River mainstem from Bonneville Dam at Rkm 235, west to confluence of the Columbia River with the Pacific Ocean. It is comprised of six 4th field HUCs ranging in size from 1,753−3,756 km2 (Table 1). Watersheds within the Lower Columbia River sub-unit include the Lower Columbia-Sandy, Lewis, Upper and Lower Cowlitz, Lower Columbia-Clatskanie, and Lower Columbia River (Figure 1). Figure 1. Map of watersheds within the Lower Columbia/Willamette RMU, Lower Columbia sub-unit. 1 Lower Columbia sub-unit – Regional Implementation Plan August 6, 2018 Table 1. Drainage Size and Level III Ecoregions of the 4th Field Hydrologic Unit Code (HUC) Watersheds located within the Lower Columbia sub-unit. HUC Drainage Watershed Number Size (km2) Level III Ecoregion(s) Lower Columbia-Sandy 17080001 2,263 Willamette Valley, Cascades Lewis 17080002 2,719 Puget Lowland, Willamette Valley, Cascades Upper Cowlitz 17080004 2,654 Puget Lowland Lower Cowlitz 17080005 3,756 Puget Lowland, Cascades Lower Columbia-Clatskanie 17080003 2,349 Coast Range, Willamette Valley Lower Columbia 17080006 1,753 Coast Range B. Status of Species Conservation Assessment and New Updates Current Pacific Lamprey distribution in the Lower Columbia sub-unit is greatly reduced from historical range (Table 2). -

Oregon Native Fish Status Report – Volume II
Oregon Native Fish Status Report – Volume II Chum 211 Oregon Native Fish Status Report – Volume II Coastal Chum Existing Populations Chum salmon found along the Oregon Coast are at the southern end of the species’ range. There is limited information on the historic distribution of chum salmon, which has made it difficult to determine how many historic populations there were and where they were located. The identification of thirteen historical populations was based on historical records of commercial landings of chum from bay and river fisheries (Table 80). Only occasional records were found to substantiate the existence of the five most southern populations. In the Alsea and Coos basins, there continue to be occasional sightings of chum salmon during fall chinook and coho spawning surveys. It is possible the occasional historic landings and current sightings in the southern basins were the result of non-native chum entering these bays or misidentification. For this report, we have identified the five southern-most populations as either extinct or presumed extinct (see “Population Details” below). A more thorough examination of the evidence during the development of a conservation plan for the Coastal Chum SMU may determine that there were no historical populations south of the Yaquina River. Effects of historical fisheries on existing populations are unclear but it is possible that the current distribution and status of populations has been shaped by historical fisheries. Table 80. Population list and existence status for the Coastal Chum SMU. Exist Population Description Yes Necanicum Necanicum River basin plus ocean tributaries south to Cape Falcon. Yes Nehalem Nehalem River basin. -

Backcountry Campsites at Waptus Lake, Alpine Lakes Wilderness
BACKCOUNTRY CAMPSITES AT WAPTUS LAKE, ALPINE LAKES WILDERNESS, WASHINGTON: CHANGES IN SPATIAL DISTRIBUTION, IMPACTED AREAS, AND USE OVER TIME ___________________________________________________ A Thesis Presented to The Graduate Faculty Central Washington University ___________________________________________________ In Partial Fulfillment of the Requirements for the Degree Master of Science Resource Management ___________________________________________________ by Darcy Lynn Batura May 2011 CENTRAL WASHINGTON UNIVERSITY Graduate Studies We hereby approve the thesis of Darcy Lynn Batura Candidate for the degree of Master of Science APPROVED FOR THE GRADUATE FACULTY ______________ _________________________________________ Dr. Karl Lillquist, Committee Chair ______________ _________________________________________ Dr. Anthony Gabriel ______________ _________________________________________ Dr. Thomas Cottrell ______________ _________________________________________ Resource Management Program Director ______________ _________________________________________ Dean of Graduate Studies ii ABSTRACT BACKCOUNTRY CAMPSITES AT WAPTUS LAKE, ALPINE LAKES WILDERNESS, WASHINGTON: CHANGES IN SPATIAL DISTRIBUTION, IMPACTED AREAS, AND USE OVER TIME by Darcy Lynn Batura May 2011 The Wilderness Act was created to protect backcountry resources, however; the cumulative effects of recreational impacts are adversely affecting the biophysical resource elements. Waptus Lake is located in the Alpine Lakes Wilderness, the most heavily used wilderness in Washington -

A Geophysical Study of the North Scappoose Creek, Alder Creek, Clatskanie River Lineament, Along the Trend of the Portland Hills Fault, Columbia County, Oregon
Portland State University PDXScholar Dissertations and Theses Dissertations and Theses 1982 A geophysical study of the North Scappoose Creek, Alder Creek, Clatskanie River lineament, along the trend of the Portland Hills fault, Columbia County, Oregon Nina Haas Portland State University Let us know how access to this document benefits ouy . Follow this and additional works at: http://pdxscholar.library.pdx.edu/open_access_etds Part of the Geology Commons, and the Tectonics and Structure Commons Recommended Citation Haas, Nina, "A geophysical study of the North Scappoose Creek, Alder Creek, Clatskanie River lineament, along the trend of the Portland Hills fault, Columbia County, Oregon" (1982). Dissertations and Theses. Paper 3254. 10.15760/etd.3244 This Thesis is brought to you for free and open access. It has been accepted for inclusion in Dissertations and Theses by an authorized administrator of PDXScholar. For more information, please contact [email protected]. AN ABSTRACT OF THE THESIS OF Nina Haas for the Master of Science in Geology presented December 15, 1982. Title: A Geophysical Study of the North Scappoose Creek - Alder Creek - Clatskanie River Lineament, Along the Trend of the Portland Hills Fault, Columbia County, Oregon. APPROVED BY MEMBERS OF THE THESIS COMMITTEE: Chairman Giibett • Benson The Portland Hills fault forms a strong northwest trending lineament along the east side of the Tualatin Mountains. An en echelon lineament follows North Scappoose Creek, Alder Creek, and the Clatskanie River along the same trend, through Columbia County, Oregon. The possibility that this lineament follows a fault or fault zone was investigated in this study. Geophysical methods were used, with seismic 2 refraction, magnetic and gravity lines run perpendicular to the lineament. -

Sea-Level Rise for the Coasts of California, Oregon, and Washington: Past, Present, and Future
Sea-Level Rise for the Coasts of California, Oregon, and Washington: Past, Present, and Future As more and more states are incorporating projections of sea-level rise into coastal planning efforts, the states of California, Oregon, and Washington asked the National Research Council to project sea-level rise along their coasts for the years 2030, 2050, and 2100, taking into account the many factors that affect sea-level rise on a local scale. The projections show a sharp distinction at Cape Mendocino in northern California. South of that point, sea-level rise is expected to be very close to global projections; north of that point, sea-level rise is projected to be less than global projections because seismic strain is pushing the land upward. ny significant sea-level In compliance with a rise will pose enor- 2008 executive order, mous risks to the California state agencies have A been incorporating projec- valuable infrastructure, devel- opment, and wetlands that line tions of sea-level rise into much of the 1,600 mile shore- their coastal planning. This line of California, Oregon, and study provides the first Washington. For example, in comprehensive regional San Francisco Bay, two inter- projections of the changes in national airports, the ports of sea level expected in San Francisco and Oakland, a California, Oregon, and naval air station, freeways, Washington. housing developments, and sports stadiums have been Global Sea-Level Rise built on fill that raised the land Following a few thousand level only a few feet above the years of relative stability, highest tides. The San Francisco International Airport (center) global sea level has been Sea-level change is linked and surrounding areas will begin to flood with as rising since the late 19th or to changes in the Earth’s little as 40 cm (16 inches) of sea-level rise, a early 20th century, when climate. -

A Brief History of the Umatilla National Forest
A BRIEFHISTORYOFTHE UMATILLA NATIONAL FOREST1 Compiled By David C. Powell June 2008 1804-1806 The Lewis and Clark Expedition ventured close to the north and west sides of the Umatilla National Forest as they traveled along the Snake and Columbia rivers. As the Lewis & Clark party drew closer to the Walla Walla River on their return trip in 1806, their journal entries note the absence of firewood, Indian use of shrubs for fuel, abundant roots for human consumption, and good availability of grass for horses. Writing some dis- tance up the Walla Walla River, William Clark noted that “great portions of these bottoms has been latterly burnt which has entirely destroyed the timbered growth” (Robbins 1997). 1810-1840 This 3-decade period was a period of exploration and use by trappers, missionaries, natu- ralists, and government scientists or explorers. William Price Hunt (fur trader), John Kirk Townsend (naturalist), Peter Skene Ogden (trap- per and guide), Thomas Nuttall (botanist), Reverend Samuel Parker (missionary), Marcus and Narcissa Whitman (missionaries), Henry and Eliza Spaulding (missionaries), Captain Benjamin Bonneville (military explorer), Captain John Charles Fremont (military scientist), Nathaniel J. Wyeth (fur trader), and Jason Lee (missionary) are just a few of the people who visited and described the Blue Mountains during this era. 1840-1859 During the 1840s and 1850s – the Oregon Trail era – much overland migration occurred as settlers passed through the Blue Mountains on their way to the Willamette Valley (the Oregon Trail continued to receive fairly heavy use until well into the late 1870s). The Ore- gon Trail traversed the Umatilla National Forest. -

Umatilla National Forest 2019 Personal-Use Firewood Maps Attachment–Part 2 (Part 1 Is Your Permit Form)
United States Department of Agriculture Umatilla National Forest 2019 Personal-Use Firewood Maps Attachment–Part 2 (Part 1 is your Permit Form) Is Today a Cut Day? INSIDE......... It's Your Responsibility to Important News for 2019.................................2 Find Out Before You Head Out! Heppner District Maps................................5-6 An updated recorded message will let you know if firewood North Fork John Day District Maps.…..............6-7 cutting is allowed, restricted to certain times of the day, or Walla Walla District Maps.............................8-10 closed completely due to hot, dry weather conditions. Pomeroy District Maps...……..….......…....11-12 21" Ruler for gauging diameter............................8-9 Call Toll-Free 2019 Firewood Season Calendar……...….….13 1-877-958-9663 Where to call for information .…......................16 Page 2 Umatilla National Forest's 2019 Program GENERAL INFORMATION: COMMERCIAL FIREWOOD: To purchase a firewood permit, you must be 18 years of age or older and All commercial activities on National Forest System Lands require a present a government-issued photo ID. commercial permit. If you wish to cut and sell firewood commercially, you must purchase a commercial firewood permit through the local The minimum cost for a personal-use firewood permit is $20, which buys Ranger District office for your area of interest. District contact four-cords. Anything over four cords will cost an additional $5 per cord. information is provided on the back page of this guide. Each household is allowed a maximum limit of 12 cords per year. HEPPNER DISTRICT OFFERS LIVE JUNIPER CUTTING: Firewood permits are available at all Umatilla National Forest Offices and at several local vendors. -

Witd Attd Scettic "Ri()E1t Stadr "Repo1tt
Ri.;-~, vev.:i A-':> l/1711at'U'"vr /J<.va AerJl../G.. /IS'iii·r. 1/1;([~~c":.b;f.. (i..C../J,~T!vt,~l.{·s NG~ oF r.Jv1,.,r I~ 1'?'7$ THE /2£;; 'te.t ~IU { 0 /~yu,,,/:--(,/tit,.J> J~<t 1L10 Witd attd Scettic "Ri()e1t Stadr "Repo1tt TABLE OF CONTENTS PART I - SUMMATION CHAPTER 1 - INTRODUCTION THE WILD AND SCENIC RIVERS ACT ............................................2 AGENCY INTERPRETATION ...................................................2 STUDY RIVERS ..............................................................2 STUDY APPROACH ..........................................................4 CHAPTER2-SUMMARYOF FINDINGS & RECOMMENDATIONS ......................... FINDINGS ..................................................................4 RECOMMENDATIONS ........................................................5 PART II - THE STUDY CHAPTER 1 - INVENTORY .......................................................... -8 THE DRAINAGE BASIN .......................................................8 THE STUDY AREA .......................................................... · 10 CHAPTER2-EVALUATION OF RIVERS &ADJOINING LANDS CRITERIA USED ............................................................ 13 ANALYSIS ................................................................. 15 OUTSTANDING CHARACTERISTICS ........................................... 20 CHAPTER 3 -CLASSIFICATION ALTERNATIVES, ANALYSIS & SELECTION OF STUDY PROPOSAL BASIS OF ANALYSIS ........................................................ 23 THE PRESENT SITUATION .................................................. -

Page 1 of 2 Colville National Forest
Colville National Forest - Salmo-Priest Wilderness Page 1 of 2 United States Department of Agriculture Colville National Forest Forest Service Forest Service Home About the Agency Contact the National Office Search Go Salmo-Priest Wilderness Site Map Area Status: Open Colville National Forest Home Tucked among the Selkirk Mountains in the extreme northeastern corner of Washington, the U-shaped 43,348 Special Places Low Fire Danger acre Salmo-Priest Wilderness extends its borders along those of Idaho and British Columbia. Its most prominent Recreation Quick Links features are two very long ridges, generally running Bicycling southwest to northeast, connected near their northern Salmo-Priest Wilderness Camping & Cabins ends by a ridge crowned by 6,828-foot Salmo Mountain. Winter Recreation in Washington State Climbing The eastern ridge stands lower, more wooded, more rounded off and more Upper Columbia Children's Forest Fishing accessible than the steep-sided, rocky-crested western ridge. Streams have cut Hiking deep drainages into both ridges. Water from the eastern side of the eastern ridge ends up in Idaho's Priest River. The remaining wilderness drains generally westerly Areas & Activities Horse Riding & Camping via Sullivan Creek and the Salmo River into the Pend Oreille River. Find An Area Hunting Below the ridge tops of this well-watered Wilderness (at 50+ inches of precipitation Highlights Nature Viewing annually) you'll find the largest growth of virgin forest left in eastern Washington: western red cedar, western hemlock, Douglas fir, grand fir, larch. The forest houses Recreation Passes OHV Riding & Camping mule deer and white-tailed deer, elk, black bears, cougars, bobcats, badgers, pine Recreation Passes martens, lynx, bighorn sheep, and moose. -

Heaven in Hell's Canyon
Northwest Explorer ONDAL ONDAL M M EN EN K K Left: Hiker at the boundary of Hell’s Canyon Wilderness. Right: Approaching Horse Heaven, elevation 8,100 feet on the Seven Devils Loop Trail in Hell’s Canyon Wilderness. June and July are good times to explore Washington’s southeast corner in the Wenaha-Tucannon area and in the nearby Hells Canyon area of Oregon and Idaho. Heaven in Hell’s Canyon Hiking two wilderness areas near Washington’s southeast corner By Ken Mondal finding places to backpack when the high jaw-dropping views. This hike can be country is snowed in. The Imnaha River done comfortably in 3-4 days. Seven Devils Loop in Hell’s is hikable virtually year round. An excellent description of hiking the Canyon is Heavenly On the Idaho side of the recreation Seven Devils Loop can be found in Hik- There is no question that the Grand area is a 215,000-acre wilderness area, ing Idaho by Maughan and Maughan, Canyon is one of the natural wonders which includes the Seven Devils Moun- published by Falcon. For the other hikes of the world. However, if one measures tains. The premier hike within this I would recommend Rich Landers’ 100 from the Snake River to the summit of wilderness is the Seven Devils Loop, a Hikes in the Inland Northwest. 9,393 foot He Devil Peak in the Seven rugged 29-mile round trip offering mag- Devils Mountains, this makes Hells nificent views into Hells Canyon many Choose Forgotten Wenaha- Canyon the deepest canyon in North thousands of feet below and equally Tucannon for Solitude America. -

Shedroofdivide.Pdf
Access: From Hwy 31 head east over the Ione Bridge 13 miles past Shedroof Divide #512, Salmo Basin #506, Thunder Cr Sullivan Lake. Turn right on Sullivan Cr. Rd. Travel 6 miles. Stay # 526, Jackson Cr #311, Hughes Fork # 312 left and travel 6 miles to the Thunder Cr Trail #526. Travel .5 miles Characteristics #512 #506 #526 #311 #312 and turn left on Forest Rd 260. Travel 6.5 miles to Salmo Priest Trail- head #506. Elevation at Trailhead 5450 ft 5920 ft 4750 ft 3000 ft 2970 ft Highest Point 6500 ft 6420 ft 5550 ft 5520 ft 5510 ft From Hwy 31 head east over the Ione Bridge 13 miles past Elevation Change 1550 ft 470 ft 800 ft 2520 ft 2540 ft Sullivan Lake. Turn right on Sullivan Cr. Rd. Travel 6 miles. Stay Length 22 Mi. 12 Mi. 5 Mi. 4 Mi. 6.4 Mi. right. Travel 7.8 miles on Sullivan/Pass Cr Rd to the Shedroof Trail- Difficulty Difficult Difficult Difficult Difficult Difficult head #512. The trail takes off from the north side of the road. Travel a short distance further east for parking. Season Summer Summer Summer Summer Summer v Points of Interest Description: This complex of trails provides access into and through the east arm of the Salmo-Priest Wilderness Area. These are difficult but spec- tacular hikes. Summer brings a profusion of flowers and berries but also Grizzly Bears. These trails are only lightly used. Check in View east from Shedroof Trailhead Pass Creek Pass at the Sullivan Lake Ranger Station for trail History: Shedroof is the highest and longest wilder- Cautions: closures and other ness trail in the Salmo-Priest Wilderness. -
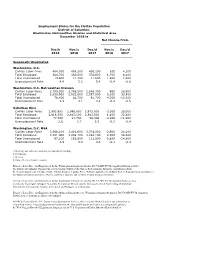
Employment Status for the Civilian Population District of Columbia, Washington Metropolitan Division and Statistical Area December 2018/A Net Change From
Employment Status for the Civilian Population District of Columbia, Washington Metropolitan Division and Statistical Area December 2018/a Net Change From Dec/b Nov/c Dec/d Nov/c Dec/d 2018 2018 2017 2018 2017 Seasonally Unadjusted Washington, D.C. Civilian Labor Force 404,300 404,200 400,200 100 4,100 Total Employed 384,700 383,000 378,600 1,700 6,100 Total Unemployed 19,600 21,200 21,600 -1,600 -2,000 Unemployment Rate 4.8 5.2 5.4 -0.4 -0.6 Washington, D.C. Metropolitan Division Civilian Labor Force 2,709,300 2,708,500 2,688,700 800 20,600 Total Employed 2,630,900 2,625,800 2,597,000 5,100 33,900 Total Unemployed 78,400 82,700 91,700 -4,300 -13,300 Unemployment Rate 2.9 3.1 3.4 -0.2 -0.5 Suburban Ring Civilian Labor Force 2,993,800 2,996,800 2,973,800 -3,000 20,000 Total Employed 2,916,300 2,915,100 2,883,500 1,200 32,800 Total Unemployed 77,500 81,700 90,300 -4,200 -12,800 Unemployment Rate 2.6 2.7 3.0 -0.1 -0.4 Washington, D.C. MSA Civilian Labor Force 3,398,100 3,401,000 3,374,000 -2,900 24,100 Total Employed 3,301,000 3,298,100 3,262,100 2,900 38,900 Total Unemployed 97,100 102,900 111,900 -5,800 -14,800 Unemployment Rate 2.9 3.0 3.3 -0.1 -0.4 a/ Data may not add to the totals due to independent rounding.