Reviewing Phosphorus and Nitrogen Targets for the SSSI Meres
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
Development Management Report
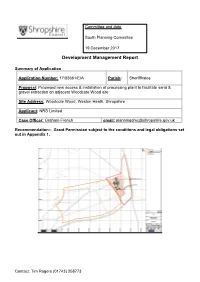
Committee and date South Planning Committee 19 December 2017 Development Management Report Summary of Application Application Number: 17/03661/EIA Parish: Sheriffhales Proposal: Proposed new access & installation of processing plant to facilitate sand & gravel extraction on adjacent Woodcote Wood site Site Address: Woodcote Wood, Weston Heath, Shropshire Applicant: NRS Limited Case Officer: Graham French email: [email protected] Recommendation:- Grant Permission subject to the conditions and legal obligations set out in Appendix 1. Contact: Tim Rogers (01743) 258773 Woodcote Wood, Weston Heath, Shropshire Planning Committee – 19 December 2017 (17/03661/EIA) REPORT 1.0 BACKGROUND 1.1 Woodcote Wood is identified as a ‘preferred area’ for sand and gravel extraction in the Shropshire Telford and Wrekin Minerals Local Plan (1996-2006). The policies of this plan are currently ‘saved’ in Telford and Wrekin and have been superseded in the Shropshire Council administrative area by the Shropshire Core Strategy and the SAMDev plan. 1.2 The planning committee of the former Shropshire County Council resolved to approve proposals to extract sand and gravel at Woodcote Wood at its meeting on 25/7/06 (ref. SC/MB2005/0336/BR). The proposals involve extraction of 2.55 million tonnes of sand and gravel at a rate of approximately 200,000 tonnes per annum, giving an operational life of some 13 years. The approval resolution was subject to a legal agreement covering off site highway matters and other issues. 1.3 The proposals involved access via a proposed new roundabout at the junction of the A41 and the Sheriffhales Road. However, the third party land required to construct the roundabout was not made available. -

Habitats Regulations Assessment
Regulation 19 Pre-Submission Draft Shropshire Local Plan 2016 to 2038 Habitats Regulations Assessment December 2020 Regulation 19 Pre-Submission Draft Shropshire Local Plan 2016-2038 : HRA Contents 1. Introduction ______________________________________________________ 4 The Habitats Regulations Assessment process __________________________________ 4 Background to the Regulation 19: Pre-Submission Draft Shropshire Local Plan 2016 – 2038 and the HRA Screening Reports _________________________________________ 5 2. Stage 1 Screening __________________________________________________ 7 Aims of the screening process and methods used _______________________________ 7 Identification of international sites requiring consideration _______________________ 7 Collation of information on the selected international sites _______________________ 9 Identifying possible mechanisms for significant effects (impact pathways) ___________ 9 Screening of international sites and potential impact pathways from policies, settlements, allocated and saved sites in the DLP _______________________ 12 Air pollution ____________________________________________________________ 12 Diffuse or regional air pollution ____________________________________________________ 13 Local sources of air pollution ______________________________________________________ 15 Hydrological impacts _____________________________________________________ 28 Recreation and other disturbance pathways __________________________________ 34 Light pollution ___________________________________________________________ -

Appendix a Environmental Baseline
Appendix A Environmental Baseline . Introduction The data collected to characterise the baseline environment of Telford and Wrekin Borough has been derived from numerous secondary sources, which are referenced as footnotes in this report. No new investigations or surveys have been undertaken. In some instances, it has been noted that different secondary sources present conflicting information and it has not been possible to verify which sources are the most accurate. Where this has been identified, the limitations have been noted. It should be noted that there is an abundance of environmental information available. However, the information presented in this Appendix has been chosen on the basis that it may be influenced or affected by the Local Flood Risk Management Strategy (LFRMS). Steps have been taken to avoid including information which is of no clear relevance to the LFRMS. It may be necessary to collect further data against which to assess the potential environmental effects of the LFRMS with regard to monitoring requirements. Population .. Population The topic of population is considered first in the baseline information, since the over-arching purpose of the LFRMS is to reduce flood risk to people and property. The LFRMS also seeks to increase public awareness of flooding and promote individual and community level flood resilience. A number of properties in the Telford and Wrekin Area are in areas at risk of flooding and were affected by flooding during the Summer 2007 floods. Some of the properties were affected by flooding from fluvial sources (streams, rivers) but many properties were affected from surface water flooding from sewers and drainsi. -

Late Holocene Environmental Change at Aqualate Mere Staffordshire, UK Ledger, P
Late Holocene environmental change at Aqualate Mere Staffordshire, UK Ledger, P. Submitted version deposited in CURVE April 2011 Original citation: Ledger, P. (2010). Late Holocene environmental change at Aqualate Mere Staffordshire, UK. Unpublished MSc by Research Thesis. Coventry: Coventry University. Copyright © and Moral Rights are retained by the author. A copy can be downloaded for personal non-commercial research or study, without prior permission or charge. This item cannot be reproduced or quoted extensively from without first obtaining permission in writing from the copyright holder(s). The content must not be changed in any way or sold commercially in any format or medium without the formal permission of the copyright holders. One map has been removed for copyright reasons. The unabridged version of the thesis can be viewed at the Lanchester Library, Coventry University. CURVE is the Institutional Repository for Coventry University http://curve.coventry.ac.uk/open 0 Late Holocene Environmental Change at Aqualate Mere, Staffordshire, U.K. P. M. Ledger A thesis submitted in partial fulfilment of the University’s requirements for the Degree of Master of Research 2010 Coventry University 1 Abstract Palaeoecologial reconstruction of the last 3000 years was undertaken on a core from Aqualate Mere, Staffordshire, U.K. Diatom analysis was the main tool used for reconstruction and the data were analysed in conjunction with previously acquired pollen and environmental magnetics data. Core chronology is provided by 14C and SCP measurements, also taken from previous work by Pittam (2006). The findings of the investigation point to a mixture of direct climatic forcing, human mediated climatic forcing, and direct anthropogenic forcing on palaeoecology. -

Shropshire Middle Severn Abstraction Licensing Strategy
Shropshire Middle Severn Abstraction Licensing Strategy A strategy to manage water resources sustainably July 2021 Page 1 of 41 We are the Environment Agency. We protect and improve the environment. We help people and wildlife adapt to climate change and reduce its impacts, including flooding, drought, sea level rise and coastal erosion. We improve the quality of our water, land and air by tackling pollution. We work with businesses to help them comply with environmental regulations. A healthy and diverse environment enhances people's lives and contributes to economic growth. We can’t do this alone. We work as part of the Defra group (Department for Environment, Food & Rural Affairs), with the rest of government, local councils, businesses, civil society groups and local communities to create a better place for people and wildlife. Published by: Environment Agency Horizon House, Deanery Road, Bristol BS1 5AH www.gov.uk/environment-agency © Environment Agency 2021 All rights reserved. This document may be reproduced with prior permission of the Environment Agency. Further copies of this report are available from our publications catalogue: http://www.gov.uk/government/publications or our National Customer Contact Centre: 03708 506 506 Email: [email protected] 2 of 41 Contents 1. About the licensing strategy ...................................................................................... 4 1.1. Overview ................................................................................................................ -

M E R E S a N D M O S S E S C O N S E R V a T I O N P L a N S R O S T H E R N E M E R E Report To: Prepared By: English Nature (
Meres and mosses conservation plan: Rostherne Mere Item Type monograph Publisher Environmental Consultancy University of Sheffield Download date 30/09/2021 23:32:19 Link to Item http://hdl.handle.net/1834/27264 M eres and M osses C onservation Plans R ostherne M ere Report to: Prepared by: English Nature (West Midlands Team), Attingham Park, Shrewsbury, Environmental Consultancy Shropshire, SY4 4TW University of Sheffield .environment Agency (North-West Region), PO Box 12, Richard Fairclough House, Endcliffe Holt Nutsford Road, Warrington, Cheshire WA4 1HG 343 Fulwood Road environment Agency (Midlands Region), Hafren House, Welshpool Road, Sheffield S10 3BQ Shelton, Shrewsbury, Shropshire SY3 8BB TEL: 0 1 1 4 2 6 6 9292 FAX: 0114 2667707 February 2001 Meres and Mosses Conservation Plans Introduction The Meres and Mosses are an internationally-important series of open water and peatland sites in the north-west Midlands. A large proportion of them are notified as Sites of Special Scientific Interest (SSSI), and more than thirty are designated as Wetlands of International Importance under the Ramsar Convention. A small number are candidate Special Areas of Conservation (SAC) under the European Habitats Directive. The characteristic habitats of the Meres and Mosses include open water lakes, reed-swamp, herbaceous fen, can woodland, wet pasture, transition mire and raised bog. In November 1998, English Nature launched a “Strategy for the conservation of the Meres and Mosses of Cheshire, Shropshire and Staffordshire”. However, it was recognised that a major constraint in the implementation of this was the lack of knowledge of the detailed hydrology and management of the catchments of these wetland areas and of the issues affecting them. -

Local Environment Agency Plan
I LeTVf^ - &OX 4 local environment agency plan MIDDLE SEVERN Second Annual Review January 2000 Marke) Draytoo OSWE Wem Ruyton xRlbwns ewport; SHREWSBi TELFORD BRIDONORTH Church Stretton KIDDERMINSTER Bewdl( Dtoitwich WORCESTER Middle Severn Local Environment Agency Plan (LEAP) Previous LEAP Documents Middle Severn LEAP Consultation Report April 1997 (now out o f stock) Middle Severn LEAP Consultation Report Summary April 1997 Middle Severn LEAP Action Plan November 1997 Middle Severn LEAP First Annual Review January 1999 This document should be read in conjunction with the Middle Severn LEAP Action Plan. If you or your organisation need further information or copies of any of the above documents, please contact: Jo Langfield Acting LEAPs Team Leader Environment Agency Hafren House Welshpool Road Shelton Shrewsbury Shropshire SY3 8BB Telephone: (01743) 272828 Fax: (01743) 272138 E-mai 1: jo. [email protected]. uk MI - 01/00 - 400 - C - BEXN FOREWORD The Environment Agency was established to protect, monitor and improve the environment in its broadest sense - ultimately contributing to the worldwide goal of sustainable development. One way of achieving this aim is through the production of Local Environment Agency Plans (LEAPs), which provide a framework for protecting and improving our local environment. This is the second Annual Review of the Middle Severn LEAP. Its purpose is to highlight the progress that has been made by the Agency and others during the past year, in tackling the issues and problems identified in the Action Plan. The progress that has been made over the last twelve months reflects the commitment of all those involved in the plan, and highlights the importance of the partnership approach developed through the LEAP process. -

Local Environment Agency Plan
local environment agency plan WEAVER/DANE CONSULTATION REPORT OCTOBER 1997 nutsford Biddul, E n v ir o n m e n t A g e n c y NATIONAL LIBRARY & INFORMATION SERVICE ANGLIAN REGION Kingfisher House, Goldhay Way, Orton Goldhay, Peterborough PE2 5ZR En v ir o n m e n t Ag ency Weaver/Dane Key Details • Total Area 1423 km2 • Population c.500,000 • Administrative Details District Councils Vale Royal Chester Newcastle-under-Lyme Congleton High Peak Warrington North Shropshire Crewe & Nantwich Staffordshire Moorlands Halton Macclesfield • Water Resources Largest Abstraction (at Brunner Mond, Northwich) 165,478 m3/day Average Annual Rainfall 716 mm Number of River Level only Measuring Stations 3 Number of River Level & Flow Measuring Stations 7 Number of Raingauges 25 • Flood Protection Length of Designated "Main River" Watercourses 619.4 km (Maintained by the Environment Agency) • Water Quality Length of classified river and canal 623.8 km • Fisheries Length of trout fishery 93 km Length of coarse fishery 187 km • Conservation Number of Sites of Special Scientific Importance (SSSI) 81 Number of Sites of Biological Importance (SBI) 412 • Heritage Sites Number of Scheduled Ancient Monuments (SAMs) 30 Number of Conservation Areas 12 NB These figures are for designated sites located near to "main river" & therefore, do not include all sites within the area boundary. • Integrated Pollution Control/Radioactive Substances I PC sites 21 Authorised Processes 49 RAS Authorisations 7 Registrations 33 • Waste Regulation Number of licensed sites 63 Number of registered exempt sites 114 Number of registered carriers of waste 375 ENVIRONMENT AGENCY Weaver/Dane LEAP The Environment Agency Kingfisher House, Goldhay Way, Orton Goldhay, i ^ ENVIRONMENT Peterborough PE2 5ZR. -

A Photographic Journey Along the River Tern by Andrew D. Boden
Tern Views A photographic journey along the river Tern by Andrew D. Boden Contents Introduction and acknowledgements Pages ii & iii List of illustrations Pages iv & v Map 1 Part 1 - From Maer to Market Drayton Pages 1 to 11 Maps 2 & 3 Part 2 - From Market Drayton to Crudgington Pages 12 to 22 Map 4 Part 3 - From Crudgington to the Severn Pages 23 to 31 Appendix I - Domesday Book References Pages vi & vii Appendix II - Books and other reference material Page viii Appendix III - The Tern in Flood October 1998 Page ix Front Cover - the Tern at Upton Forge 1998 i Introduction This short book began as a few photographs for my wife’s class of Market Drayton schoolchil- dren, but as I began to follow the Tern from its source to its eventual meeting with the Severn I found out many interesting facts and talked to numerous pleasant and helpful people, all of whom were enthusiastic about their particular stretch of the river and most willing to assist me with facts, stories and reminiscences. The abundance and variety of flora and fauna was also an interesting aspect of the changing environment along the river; indeed it became difficult to decide what to include and what to omit. I therefore decided that the place of this book was to provide an intro- duction to the river as it is at present, with references to its past life and work, leaving any deeper research into particular features to those with more particular or specialist interest in specific areas and buildings. -

Chester Zoo Nature Reserve News by Andy Jennings-Giles 6 - 10
Issue 1 Winter 2016 The Local Environmental Records centre serving Cheshire, Halton, Warrington and Wirral Issue 2 Spring 2017 1 Hoverfly (Eristalis sp.) Contents RECORD Round up 3 Wildlife on your doorstep by Lisa Reeves 4 - 5 Chester Zoo Nature Reserve News by Andy Jennings-Giles 6 - 10 Snap That Fly! by Phil Brighton 11 - 13 Cheshire’s Meres by Katie Piercy 13 - 14 Count Me in Graduates 15 - 16 Species Spotlight - Swifts by Susie Phillips 17 - 18 What to look out for in the coming months 18 - 20 Upcoming events in Cheshire 21 - 28 A note from the editor… Welcome to the Spring 2017 issue of RECORD’s newsletter. Thank you again to everyone who has contributed. We have lots of interesting articles from our volunteers, partners and recorders that are sure to inspire everyone to get out recording this spring. We also have an insightful up- date about the development of Chester Zoo’s own nature reserve. At the end of the newsletter you will also find details of the upcoming RECORD Conference, be sure to sign up and reserve your place to join us on the 7th May. Please get in touch if you would like to contribute to our Summer 2017 newsletter. Leanna Dixon (Data Services Officer, [email protected]) 2 RECORD Round Up Despite the cold weather over the winter months there has still been lots going on at RECORD. Our volunteers have been very busy inputting wildlife rec- ords onto our system with over 23,000 new records add- ed this winter and a total of 52,375 over the last year! Over 52,000 records In the last month we have also run two wildlife recording have been input by our workshops. -

Screening Report
Shropshire Council Local Plan Review 2016 - 2036 Preferred Options: Scale and Distribution of Development Habitats Regulations Assessment Screening Report October 2017 LPR Preferred Options:Scale and Distribution of Development HRA Screening Report Contents 1. Introduction _____________________________________________________ 3 What are Habitats Regulations Assessments? ____________________________ 3 Background to the Shropshire Local Plan Review 2016-2036 and the HRA Screening Reports _________________________________________________ 4 2. Methodology ____________________________________________________ 6 Purpose of the HRA Screening Report __________________________________ 6 Identification of international sites requiring consideration ___________________ 6 Collation of information on the above international sites ____________________ 7 Identifying possible mechanisms for significant effects (effect pathways) _______ 8 Table 1: General effect pathways ______________________________________ 9 3. Screening or assessment of potential effects _____________________ 10 Air pollution ______________________________________________________ 10 Water pathways __________________________________________________ 11 Recreation pathways: ______________________________________________ 12 Screening of policy options __________________________________________ 12 Summary of the results of initial HRA screening __________________________ 13 4. In-combination effects __________________________________________ 14 5. Conclusions and recommendations ______________________________ -

Woore Neighbourhood Plan 2016-2036 - HRA Screening Report
Woore Neighbourhood Plan 2016-2036 - HRA Screening Report Woore Neighbourhood Plan 2016-2036 Regulation 15 Submission Version May 2018 Habitats Regulations Assessment Screening Report Updated October 2018 Updated October 2018 1 Woore Neighbourhood Plan 2016-2036 - HRA Screening Report Contents Page 1 Introduction 4 1.1 What are Habitats Regulations 4 Assessments? 1.2 Background to the WNP 2016-2036 and the 5 HRA Screening Report 2 Methodology 6 2.1 Purpose of the HRA Screening Report 6 2.2 Identification of international sites requiring 7 consideration 2.3 Collation of information on international 7 sites 2.4 Identifying possible mechanisms for 8 significant effects (effect pathways) 2.5 Detailed consideration of potential effect 9 pathways and buffer distances where applicable 2.5.1 Air pollution 9 2.5.2 Hydrological impacts (water pathways) 10 2.5.3 Recreation pathways 11 2.5.4 Biosecurity pathways 12 2.5.5 Lighting pathways 12 3 Screening or assessment of potential effects 3.1 Initial screening of policies and site 13 allocations in the WNP 3.2 Screening of likely significant effects on 13 international sites via the remaining policy in the WNP 4 Conclusions and 24 recommendations 5 Public consultation 24 6 References and 24 abbreviations Appendix 1 Maps of international sites considered in this 27 report Appendix 2 Description of international sites including 37 conservation objectives and sensitivities Appendix 3 Screening of WNP policies for potential 58 significant effects Tables Page Table 1 General effect pathways 8 Updated October 2018 2 Woore Neighbourhood Plan 2016-2036 - HRA Screening Report Table 2 Screening of Betley Mere 15 Table 3 Screening of Black Firs and Cranberry Bog 16 Table 4 Screening of Brown Moss 17 Table 5 Screening of Cop Mere 18 Table 6 Screening of Oakhanger Moss 19 Table 7 Screening of Oss Mere 20 Table 8 Screening of Wybunbury Moss 21 Table 9 Screening of River Severn 22 Updated October 2018 3 Woore Neighbourhood Plan 2016-2036 - HRA Screening Report 1.