Maestro En Ciencias Biológicas
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

A Simb Lica Do Espa O 1 .Pdf
A S I M B Ó L I C A DO ESPAÇO EM O SENHOR DOS ANÉIS DE J.R.R. TOLKIEN MARIA DO ROSÁRIO MONTEIRO LIVROS DE AREIA T Í T U L O : A simbólica do espaço em “O Senhor dos Anéis” de J. R. R. Tolkien A uto R A : Maria do Rosário Monteiro Esta edição © 2010 Livros de Areia Editores, Lda. Textos © 2010 Maria do Rosário Monteiro Desenhos de J.R.R. Tolkien © The J.R.R. Tolkien Estate Limited Reprodução autorizada por HarperCollins Publishers Ltd. Na contracapa: quadro de Caspar David Friedrich (1824) REVISÃ O D E T EX to : Piedade Pires DESI G N : Pedro Marques / Livros de Areia I MP RESSÃ O : Publidisa Texto composto em Sabon, 10 /12 pt. Títulos compostos em Albertus (fonte desenhada por Berthold Wolpe em 1932) IS B N : 978-989-81181-1-0 DE P ÓSI to L E G A L : 1 . ª E D I Ç Ã O : Novembro de 2010 Reservados todos os direitos. Esta edição não pode ser reproduzida, nem trans mitida, no todo ou em parte, por qualquer processo electrónico, mecânico, gra vação ou outros, sem prévia autorização escrita da Editora. [email protected] www.livrosdeareia.com PROJECTO APOIADO POR Grupo Centro de Interdisciplinar História de Estudos da Cultura Portugueses (Universidade Nova de Lisboa) ÍNDICE abreviaturas ............................................................................. 9 a autora ..................................................................................... 11 1. introdução ........................................................................................... 13 notas ......................................................................................................... -
Book Title Author Reading Level Approx. Grade Level
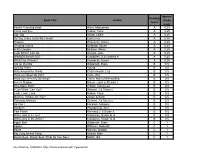
Approx. Reading Book Title Author Grade Level Level Anno's Counting Book Anno, Mitsumasa A 0.25 Count and See Hoban, Tana A 0.25 Dig, Dig Wood, Leslie A 0.25 Do You Want To Be My Friend? Carle, Eric A 0.25 Flowers Hoenecke, Karen A 0.25 Growing Colors McMillan, Bruce A 0.25 In My Garden McLean, Moria A 0.25 Look What I Can Do Aruego, Jose A 0.25 What Do Insects Do? Canizares, S.& Chanko,P A 0.25 What Has Wheels? Hoenecke, Karen A 0.25 Cat on the Mat Wildsmith, Brain B 0.5 Getting There Young B 0.5 Hats Around the World Charlesworth, Liza B 0.5 Have you Seen My Cat? Carle, Eric B 0.5 Have you seen my Duckling? Tafuri, Nancy/Greenwillow B 0.5 Here's Skipper Salem, Llynn & Stewart,J B 0.5 How Many Fish? Cohen, Caron Lee B 0.5 I Can Write, Can You? Stewart, J & Salem,L B 0.5 Look, Look, Look Hoban, Tana B 0.5 Mommy, Where are You? Ziefert & Boon B 0.5 Runaway Monkey Stewart, J & Salem,L B 0.5 So Can I Facklam, Margery B 0.5 Sunburn Prokopchak, Ann B 0.5 Two Points Kennedy,J. & Eaton,A B 0.5 Who Lives in a Tree? Canizares, Susan et al B 0.5 Who Lives in the Arctic? Canizares, Susan et al B 0.5 Apple Bird Wildsmith, Brain C 1 Apples Williams, Deborah C 1 Bears Kalman, Bobbie C 1 Big Long Animal Song Artwell, Mike C 1 Brown Bear, Brown Bear What Do You See? Martin, Bill C 1 Found online, 7/20/2012, http://home.comcast.net/~ngiansante/ Approx. -

Caso Carrà» Le Idee Delle Donne Per Un Diverso Svili
Anno 61* N. 51 Quotidiano LIRE 500 Spcd. abb. post, gruppo 1/70 * SABATO 3 MARZO 1984 Arretrati L. 1.000 I Unità ORGANO DEL PARTITO COMUNISTA ITALIANO La spinta unitaria dei lavoratori pesa sul confronto tra i sindacati Trentin: la CGIL La lotta nel Sud può e deve guidare Una folla immensa il movimento in corteo a Taranto Dalla svolta dell'Eur al decreto del 14 febbraio - Oggi l'assem Ancora adesioni alla proposta della Breda e della Falck per la blea dei quadri socialisti a Roma - Rinnovatori e conservatori manifestazione a Roma il 24 - Sì dall'assemblea di Bologna ROMA — È iniziata la batta di lavoro, subordinando a ciò il rappresentante del lavora Taranto ha vissuto ieri una straordinaria ha predisposto migliala di volantini che in glia parlamentare sul decre la stessa quantità delle ri tori in cassa integrazione. La giornata di lotta. I lavoratori confluiti in due questi giorni vengono diffusi In tutte le fab to governativo che taglia la vendicazioni salariali». logica della contrattazione cortei nel centro della città erano decine di briche. Ieri a Milano i delegati della Breda scala mobile, accompagnala — Quali altri condizioni e- centralizzata ha addirittura migliaia, non meno di 40 mila secondo le va Fucine e della Falck Concordia, e tra questi da un ampio movimento di rano necessarie per fare del accentuato le divaricazioni». lutazioni più equilibrate. È stata una rispo numerosi aderenti alla CISL, hanno illustra Palazzo Chigi lotta. Nel sindacato ti dibat sindacato un soggetto poli — E ha accentuato le diffi sta di popolo venuta da una città che è un po' to in una conferenza stampa caratteri e mo tito viene schematicamente tico? colta relative alla democra la capitale industriale di un Mezzogiorno che dalità dell'Iniziativa della quale sono stati i tradotto in una specie di •Bisognava essere in gra zia interna? questa volta non è rimasto indietro. -

Craig Wilson
ISSUE 5 THE MYTH OF REVOLT Do you sense it—on the streets, in the dreamtime, by the shore? Like a great hand that glows in the dark, groping to strangle destiny, the myth of revolt is feeling its way into existence. Let’s go searching to assemble the scattered limbs of the black god. As a start, we propose three Domains of Revolt that will act as surrealist vehicles for our summoning. Published by Peculiar Mormyrid peculiarmormyrid.com All rights reserved. No part of this book may be reproduced in any form or by any electronic or mechanical means, including information storage and retrieval systems, without written permission from the publisher or author. Texts Copyright © The Authors Images Copyright © The Artists Direction: Jason Abdelhadi, Casi Cline, Steven Cline, Angel Therese Dionne, Patrik Sampler First Edition 2017 CONTRIBUTORS Jason Abdelhadi Tori Lion Armando Urias Anderson Michael Löwy Daphnée Azoulay Andrew Mendez Maurizio Brancaleoni Thomas Mordant Maria Brothers Luiz Morgadinho Miguel de Carvalho Steve Morrison Steven Cline David Nadeau Casi Cline Lucy Zae Porter Dominick Coppi John Richardson Laura Corsiglia Jose Manuel Rojo David Coulter Ody Saban Angel Therese Dionne Ron Sakolsky Vicente Gutiérrez Escudero Mark Sanders Mattias Forshage Andria Matta May Schaal Javier Gálvez Gregg Simpson Jesús García Rodríguez Ethan Snyder Joël Gayraud Maurice Spira Jan Gillam RW Spryszak Guy Girard Dan Stanciu Annalynn Hammond Michael Vandelaar Janice Hathaway Allan Vilu Dale Houstman John Welson Karl Howeth Tim White Joseph Jablonski Craig Wilson Bruno Jacobs Claire Wylde Ian Kappos Stephen Kirin Collective Submissions: Megan Leach Chimaera Surrealist Group (Ottawa) Lee Levinson CAPA Collective Rik Lina House of Mysticum Neko Linda Williams Roger Farr/Sheila Nopper/Ron Sakolsky CONTENTS INQUIRY ....................6 TIM WHITE...................8 STEVEN CLINE..................15 CEPHALOPODA JASON ABDELHADI ............. -

Remates De Selectos 2016
REMATES DE SELECTOS 2016 Nacidos en el segundo semestre de 2014 87 PRODUCTOS SELECTOS A LA VENTA BOLSA TOTAL A REPARTIRSE APROX. US$ 360,000.00 Miércoles 1 y Martes 7 de junio de 2016 7.00 p.m. Jueves 9 de Junio de 2016 1:00 p.m. Ring de Ventas “Adolfo Vásquez Pequeño” Hipódromo de Monterrico MR. TOM THORNBURY Una vez más, Mr. Thornbury por invitación de la Asociación de Criadores de Caballos de Carrera del Perú ha evaluado a cada uno de los productos que saldrán a subasta en las tres fechas de Selectos. Mr. Tom Thornbury es Director Asociado de Ventas en Keeneland Inc., en Kentucky, EEUU, involucrado en la crianza de caballos de carreras desde muy joven cuando comenzó en el haras de su padre y su paso como presidente del “Thoroughbred Club of American” y del “Kentucky Thoroughbred Farm managers Club” le ha valido para adquirir a través del tiempo una vasta experiencia y conocimiento sobre el PSC que le ha permitido convertirse en un experto en inspecciones. Como es sabido, Mr. Thornbury ha brindado sus conocimientos para desarrollar las ventas de “Selectos” en otros mercados como: Argentina, Brasil Chile, Sudáfrica y Perú mismo, lo que da un plus a esta nueva oportunidad que se subastan los productos Selectos y estamos seguros que esto repercutirá en beneficio para quienes logren adquirir un producto en cualquiera de las tres noches de Selectos programadas. 2 REGLAMENTO GRAN SUBASTA DE SELECTOS PRODUCTOS NACIDOS EN 2014 (segundo semestre) 1.- Todos los productos vendidos y/o nominados en estas subastas podrán participar en los Clásicos de Selectos organizados para este efecto, que se correrán en el mes de junio 2016 como sigue: Ø Clásico Selectos Potrillos, 1400 metros arena Ø Clásico Selectos Potrancas, 1400 metros arena Ø Clásico Selectos Productos, 1500 metros césped 2.- El Jockey Club del Perú aportará a la bolsa el valor de una condicional clásica para cada carrera. -

Title Author Unit1 Unit2 Unit3 Book Type Genre Famis Item # ISBN
Title Author Unit1 Unit2 Unit3 Book Type Genre Famis Item # ISBN Publisher Grades F&P Level TC Level Annotation Notes Joshua's Night Whispers Angela Johnson Necessary Books 590420445 Scholastic K-2 Owl Moon Jane Yolen Necessary Books 590420445 Scholastic K-2 A Chair For My Mother Vera B. Williams Necessary Books 590331558 Scholastic K-2 Bigmama's Donald Crews Authors as Mentors 590221213 Scholastic K-2 Meanest Thing to Say Bill Cosby Authors as Mentors 590956167 Scholastic K-2 Joshua by the Sea (Board Book) Angela Johnson Authors as Mentors 531068463 Scholastic K-2 Mama Birds, Baby Birds (Board Book) Angela Johnson Authors as Mentors 53106848X Scholastic K-2 One of Three Angela Johnson Authors as Mentors 531070611 Scholastic K-2 Rain Feet (Board Book) (Hardcover) Angela Johnson Authors as Mentors 531068498 Scholastic K-2 When I Am Old with You Angela Johnson Authors as Mentors 531070352 Scholastic K-2 The Snowy Day Ezra Jack Keats Authors as Mentors 590030310 Scholastic K-2 Make Way for Ducklings Robery McCloskey Authors as Mentors 590339494 Scholastic K-2 Appalachia Cynthia Rylant Authors as Mentors 590512587 Scholastic K-2 Alexander and the Terrible, Horrible, No Good, Very Bad Day Judith Viorst Authors as Mentors 590421441 Scholastic K-2 Launching the Writing Corduroy Don Freeman Workshop 590309072 Scholastic K-2 Apples and Pumpkins Anne Rockwell Nonfiction Writing 59045191X Scholastic K-2 Night Shift Daddy Eileen Spinelli Small Moments 439221382 Scholastic K-2 Chicken Sunday Patricia Polacco Small Moments 59046244X Scholastic K-2 Just Us Women Jeannette Franklin Caines Small Moments 590446762 Scholastic K-2 Mike Mulligan and His Steam Shovel Virginia Lee Burton The Craft of Revision 59032487X Scholastic K-2 Koala Lou Mem Fox The Craft of Revision 590994662 Scholastic K-2 Do Like Kyla Angela Johnson Necessary Books PQZ368919 Scholastic K-2 Joshua's Night Whispers Angela Johnson Necessary Books PQZ42044 Scholastic K-2 Owl Moon Jane Yolen Necessary Books PQZ42044 Scholastic K-2 A Chair For My Mother Vera B. -

VIERNES 8 Y SÁBADO 9 DE MARZO EN HARAS DON ALFREDO PRESENTACIÓN HORA 11:00
LIQUIDACIÓN TOTAL PLANTEL DE YEGUAS MADRES Y PRODUCTOS DE MESES. 138 Yeguas Madres Hijas de: Southern halo- Candy Stripes- First American Honour and Glory- Roi Normand- Seattle Slew- Equalize Broad Brush – Trempolino - Confidential Talk - Bright Again – Ghaeer Ringaro -Incurable Optimist – Roy – Lode - Slew Gin Fizz – Dodge Yagli - Nugget Point- Thunder Gulch - Robin Des Pins – Ritz -Dubai Dust -Robin Des Bois - Exile King-Burooj – Ibero - Our Emblem – Marlin - Forest Wildcat - Gulpha Gorge -Asidero- entre otros. Gravidas de: Asidero-Ecclesiastic-Southern Lawyer-Ventilo 80 Destetes hijos de: Asidero - Ecclesiastic - Gulpha Gorge - Southern Lawyer - Ventilo VIERNES 8 y SÁBADO 9 DE MARZO EN HARAS DON ALFREDO PRESENTACIÓN HORA 11:00 Subasta Oribe Remates S.R.L. Financia en 12 cuotas en dólares. [email protected] / “Sólo para créditos autorizados” www.oriberemates.com Consulte su crédito con anticipación - 29168325 RUTA 1 KM 76,500 - RINCON DEL PINO -SAN JOSE - URUGUAY TEL 43406092 - 43406077 - e-mail: [email protected] DESCUENTOS ADICIONALES COMPRANDO 2 o 3 animales 3% de descuento. 4 o 5 animales 6% de descuento. 6 o más animales 10% de descuento. El orden de salida de los animales en el remate, será el correspondiente al número del presente catálogo. OFERTAS TELEFÓNICAS: 098 004 536 098 004 537 098 004 538 099 269 725 099 408896 099 208606 ESTADÍSTICAS 1º Estadística General Las Piedras por Carreras Ganadas 1995 - 1997 - 1998 - 1999 - 2000 Maroñas 2004 - 2005 - 2006 - 2007 - 2008 - 2009 - 2010 - 2011 1º Estadística General Las Piedras por Sumas Ganadas 1994 - 1995 - 1996 - 1997 - 1998 - 1999 - 2000 - 2001 Maroñas 2004 - 2005 - 2006 - 2007 - 2008 - 2009 2010 - 2011 - 2012 1º Estadística Clásica Las Piedras 1992 - 1993 - 1994 - 1995 - 1996 - 1997 - 1998 1999 2000 - 2001 - 2002 Maroñas 2004 - 2005 - 2006 - 2007 - 2008 - 2010 - 2011 1º Estadística Las Piedras Productos de 2 años 1992 - 1993 - 1995 - 1996 - 1997 - 1998 - 2000 2001 2002 - 2003. -

VISITA Dt ALEMAN a E. U. Convencionconvención Dede Happy Houston V
LEAN j RIADREAD EL TUCSONENSETUCSONENSE EL TUCSONENSETUCSONENSE El antiguo y mat mijor periódico tn Eipa Tha Southwesti Oldest and Finatt Newspaper --- --- Aol del Suroait m printed In Spaniih, It Semi Weekly. mr iifaiH Miii publlihtd Con noticia haita I Ultimo Minuto, Arti iiPi Bif Up To Date, with aitlclet of Interest, El cvlot y Unificación Lnaitt a Inter, qua pugna por la Tucioneme It dedicated to Pan American Unity y Amlitad Pan Amorioanol and riondthip J AñoMo XXXII70,11- Volumen LXI - Neme.Número 1010 Tucson,Tucson, Arizona Aneo. - Marts,Mártes, 66 dede MayoMayo dede 19471W Números del u.a tc ENTRO EN FUNCIONESFUNCIONES ELEL NUEVO ALCALDEALCAIDE DE TUCSONTUCSON 1. EXITO DE LA VISITA Dt ALEMAN A E. U. ConvencionConvención dede Happy Houston V. NotariosHotarios en Lleno dede EnergíaEnergia LIBERTADY YABUSO!! DURE En Habla AnteAnte lala Tucson Para "El Tucsonense" mecopciRecepciónonEn WashingtonWashington A AlomanAlemán 1 La a D LigaLiga dede PaisesPaises 'Iodo el dia de ayer 5 de Libertad es una cosa que todos gusta. Mayo, fue ocupadisimo, para Es inherente en el hombre, sentirse libre y gustoso el nuevo Mayor, o Alcalde, vivirr .4en unun pais, que que confierecallare lala hbarledlibertad a todosrod. Ha f'do rotando ol éxi o de Tucson, el Hon E. T. de la visita que está actu "Happy" Houston tem aunqueaunque sea solamentesolemenieante a.e la LeY,Ley. haciendo a M desde tfotadm pra ío en lj mañana, hasta Unidos el de N"é::i las horas avanzadas de la no- co Lic. Don M;.,-.te- l iLcma.i. Las pasionespasioneshumanas aurnenastienen hen.un unlimite, hm. -

I. Comunidad De Madrid
B.O.C.M. Núm. 33 MARTES 9 DE FEBRERO DE 2010 Pág. 7 I. COMUNIDAD DE MADRID B) Autoridades y Personal Cuarto De conformidad con lo establecido en la base sexta de la Or- Consejería de Economía y Hacienda den 923/1989, de 20 de abril (BOLETÍN OFICIAL DE LA COMUNI- DAD DE MADRID de 27 de abril), modificada parcialmente por la 409 ORDEN de 27 de enero de 2010, por la que se convoca un Orden de 22 de marzo de 2002 (BOLETÍN OFICIAL DE LA COMUNI- puesto de trabajo para su provisión por el sistema de Libre DAD DE MADRID de 27 de marzo), las solicitudes para tomar par- Designación. te en esta convocatoria se presentarán en el plazo de quince días hábiles, a contar desde el siguiente a la publicación de la convo- Existiendo un puesto de trabajo vacante reservado a funcionarios catoria; en el Registro de dicha Consejería o en la forma estable- de esta Administración autonómica, y siendo necesario proceder a cida en el artículo 38.4 de la Ley 30/1992, de 26 de noviembre, de su inmediata provisión, de conformidad con lo dispuesto en el ar- Régimen Jurídico de las Administraciones Públicas y del Proce- tículo 49 de la Ley 1/1986, de 10 de abril, de la Función Pública de dimiento Administrativo Común, y se dirigirán a la Secretaría Ge- la Comunidad de Madrid, neral Técnica de la Consejería de Economía y Hacienda, según modelo regulado en la Orden 2066/1998, de 30 de julio, de la Consejería de Hacienda (BOLETÍN OFICIAL DE LA COMUNIDAD DE DISPONGO MADRID de 11 de agosto). -
THE SUNDAY JOURNAL ALBUQUERQUE, August 26, 2018 F1
THE SUNDAY JOURNAL ALBUQUERQUE, auGust 26, 2018 F1 4468619 AGUILAR JESSICA 3118 HWY 28 LAS CRUCES NOTICE OF NAMES APPEARING TO BE 4272402 AGUILAR LUIS 11000 SKYLINE RD NE #12 ABQ OWNERS OF UNCLAIMED FUNDS 4432113 AGUILAR LUIS R 1705 SAN PEDRO RD NE RIO RANCHO 4516207 AGUILAR MARIA L 8101 DELILAH RD SW ABQ Pursuant to Section 7-8A-9 NMSA 1978, notice is hereby given that the persons listed below appear to be owners of unclaimed funds. To 4211535 AGUILAR MARY PO BOX 1690 ABQ obtain information concerning these properties, address all inquiries to 4538165 AGUILAR MAURICIO 1861 CARINA CT SW RIO RANCHO the New Mexico Taxation & Revenue Department/Unclaimed Property 4406010 AGUILAR RAFAEL S 20 OSAGE DR LOS LUNAS Office, PO Box 25123, Santa Fe, NM 87504 or (505) 476-1774. The inquiry must include the name and address of the owner and the iden- 4416259 AGUILAR RAMONA 1405 ELSIE ST S MORIARTY tifying PID# shown in this notice. Claim forms will only be provided to 4405950 AGUILAR RAYMOND 12008 PHOENIX AVE NE #6C ABQ the original owner or their representative. 4493632 AGUILAR ROY 8901 JEFFERSON ST #228 ABQ 4416260 AGUILERA ALZINA 1349 BURLEY CT LAS CRUCES 4383627 AGUILERA FELIPE 4801 SAN MATEO LN NE ABQ 4351214 AGUINADA MARY 9216 COOK RANCH PL SW ABQ PID# OWNER NAME ADDRESS CITY 4351159 AGUINADAMARTINEZ REINA 11100 GIBSON BLVD SE #F13 ABQ 4590177 JAMES HENRY 3410 EASTERN AVE SE ABQ 4573492 AGUIRRE ADRIAN 3506 HERMOSILLO DR LAS CRUCES 4518208 1CLICKPHYSICS 13104 DESERT MOON PL NE ABQ 4051071 AGUIRRE ARMANDO 5422 ALDEN ST SANTA TERESA 4350654 21ST -

INDEX to California Highways and Public Works 1937-1967
VOLUME I: INDEX to California Highways and Public Works 1937-1967 Compiled by: Jill Hupp Environmental Program, Cultural Studies Office California Department of Transportation, Sacramento, June 1997 CALTRANS ENVIRONMENTAL PROGRAM CULTURAL STUDIES OFFICE 1120 N Street, M.S. 27 Sacramento, CA 95814 (916) 653-0647 JUNE 1997 Copies available from the Caltrans Publications Distribution Unit for $25.00 (includes sales tax, shipping and handling). Please make checks payable to “Caltrans,” and mail to: Caltrans Publication Unit 1900 Royal Oaks Drive Sacramento, CA 95815-3800 (916) 323-5606 We do not accept purchase orders or orders by fax To place an order using your Visa or Mastercard, please call (916) 445-3520. TABLE OF CONTENTS CALIFORNIA HIGHWAYS AND PUBLIC WORKS INDEX 1937-1967 Introduction and Acknowledgments . v “California Highways and Public Works Journal, 1912-1967: A Brief History” . vi MAIN INDEX SUBJECTS ACCIDENTS . 4 ARCHAEOLOGY & PALEONTOLOGY. 6 AWARDS . 6 BRIDGES . 7 BUDGETS & FUNDING . 23 BUILDINGS . 25 CALIFORNIA HIGHWAY COMMISSION . 26 CONVICT LABOR . 26 CULVERTS . 27 DAMS & CANALS . 27 DEPARTMENT OF PUBLIC WORKS . 29 Division of Highways . 29 DISTRICTS . 30 DOCUMENTS & RECORDS . 30 EDUCATION . 32 ENVIRONMENTAL ISSUES. 32 EQUIPMENT . 33 EROSION CONTROL . 35 Bank Protection, Methods of . 35 Drainage . 35 Embankment Stabilization . 35 Slides/Slipouts . 36 EVENTS . 36 AASHO Conventions . 36 California State Fair . 37 Visits to State . 37 WASHO Conventions. 38 FERRIES . 38 FLOODING . 38 Damage Caused By . 38 Flood Control . 39 FREEWAYS . 40 GASOLINE TAXES . 41 GRADE CROSSINGS, SEPARATIONS & INTERCHANGES . 41 HIGHWAY PLANNING & DESIGN . 47 Photogrametry . 47 HISTORIC LANDMARKS & MONUMENTS . 48 Missions of California . 48 HISTORY: PEOPLE, PLACES & HISTORIC EVENTS . -

Pdf (Boe-A-2015-1732
BOLETÍN OFICIAL DEL ESTADO Núm. 44 Viernes 20 de febrero de 2015 Sec. III. Pág. 13932 III. OTRAS DISPOSICIONES MINISTERIO DE HACIENDA Y ADMINISTRACIONES PÚBLICAS 1732 Resolución de 26 de enero de 2015, del Departamento de Gestión Tributaria de la Agencia Estatal de Administración Tributaria, por la que se publica la revocación de números de identificación fiscal. Conforme a lo dispuesto en el apartado 4 de la disposición adicional sexta de la Ley 58/2003, de 17 de diciembre, General Tributaria, modificada por el artículo quinto, apartado diecisiete, de la Ley 36/2006, de 29 de noviembre, de Medidas para la Prevención del Fraude Fiscal, se dispone la publicación de la revocación de los números de identificación fiscal que figuran en el anexo. Madrid, 26 de enero de 2015.–El Director del Departamento de Gestión Tributaria de la Agencia Estatal de Administración Tributaria, Rufino de la Rosa Cordón. cve: BOE-A-2015-1732 BOLETÍN OFICIAL DEL ESTADO Núm. 44 Viernes 20 de febrero de 2015 Sec. III. Pág. 13933 ANEXO Relación de Números de Identificación Fiscal Revocados FECHA DEL NIF RAZÓN SOCIAL REGISTRO PÚBLICO DE INSCRIPCIÓN ACUERDO B06000095 AMBROSIO MARTINEZ Y CIA SL 22/10/2014 REGISTRO MERCANTIL DE BADAJOZ A84000033 THEMA ARQUITECTURA Y GESTION SA 02/10/2014 REGISTRO MERCANTIL DE MADRID A24000093 ANTRACITAS QUIÑONES SA 07/11/2014 REGISTRO MERCANTIL DE LEON B73000085 EURO-SABHER SL 16/10/2014 REGISTRO MERCANTIL DE MURCIA A79000030 TARAX SA 26/11/2014 REGISTRO MERCANTIL DE MADRID B84000082 TRINIDAD MONZON & HIJOS SL 26/11/2014 REGISTRO MERCANTIL