A COMPARATIVE STUDY of the AZYGOS VENOUS SYSTEM in MAN, MONKEY, DOG, CAT, RAT and RABBIT by DAVID BOWSHER Department of Anatomy, University of Liverpool
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Prenatal Diagnosis of a Variant of the Azygos Venous System
Lo Verso et al. SpringerPlus (2016) 5:1334 DOI 10.1186/s40064-016-2956-0 SHORT REPORT Open Access Prenatal diagnosis of a variant of the azygos venous system Clelia Lo Verso1*, Valentina Cigna2, Gianfranca Damiani2, Laura Lo Verso1, Rossella Conti3 and Vincenzo Duca3 Abstract Background: The azygos venous system consists of the azygos vein on the right side and the hemiazygos and accessory hemiazygos on the left side. The azygos vein runs through the abdominal cavity along the right side of the vertebral bodies, in a cranial direction, passes through the diaphragm and reaches the mediastinum, where it forms the arch of the azygos which flows into the superior vena cava. Along its course, the azygos vein communicates with the intercostal veins on the right, the hemiazygos vein that collects blood from the left lower intercostal veins, and accessory hemiazygos vein that drains into the left upper intercostal veins. The last two, at the level of the seventh thoracic vertebra, unite and end in the azygos vein. The accessory hemiazygos vein is normally included in the length between T4 and T8. The embryological origin of the accessory hemiazygos vein is the result of an expansion in the direction of the cranial hemiazygos vein, which comes from the left upper sovracardinale vein (Dudiak et al. in Semin Roentgenol 24(1):47–55, 1989; Radiographics 11(2):233–246, 1991; Webb et al. in Am J Roentgenol 139(1):157–161, 1982). Findings: This case report describes a rare variant of azygos vein system identified in prenatal diagnosis and con- firmed by postnatal ultrasonography. -

Vessels and Circulation
CARDIOVASCULAR SYSTEM OUTLINE 23.1 Anatomy of Blood Vessels 684 23.1a Blood Vessel Tunics 684 23.1b Arteries 685 23.1c Capillaries 688 23 23.1d Veins 689 23.2 Blood Pressure 691 23.3 Systemic Circulation 692 Vessels and 23.3a General Arterial Flow Out of the Heart 693 23.3b General Venous Return to the Heart 693 23.3c Blood Flow Through the Head and Neck 693 23.3d Blood Flow Through the Thoracic and Abdominal Walls 697 23.3e Blood Flow Through the Thoracic Organs 700 Circulation 23.3f Blood Flow Through the Gastrointestinal Tract 701 23.3g Blood Flow Through the Posterior Abdominal Organs, Pelvis, and Perineum 705 23.3h Blood Flow Through the Upper Limb 705 23.3i Blood Flow Through the Lower Limb 709 23.4 Pulmonary Circulation 712 23.5 Review of Heart, Systemic, and Pulmonary Circulation 714 23.6 Aging and the Cardiovascular System 715 23.7 Blood Vessel Development 716 23.7a Artery Development 716 23.7b Vein Development 717 23.7c Comparison of Fetal and Postnatal Circulation 718 MODULE 9: CARDIOVASCULAR SYSTEM mck78097_ch23_683-723.indd 683 2/14/11 4:31 PM 684 Chapter Twenty-Three Vessels and Circulation lood vessels are analogous to highways—they are an efficient larger as they merge and come closer to the heart. The site where B mode of transport for oxygen, carbon dioxide, nutrients, hor- two or more arteries (or two or more veins) converge to supply the mones, and waste products to and from body tissues. The heart is same body region is called an anastomosis (ă-nas ′tō -mō′ sis; pl., the mechanical pump that propels the blood through the vessels. -

Azygos Vein System Abnormality: Case Report
Gülhane Týp Dergisi 2006; 48: 180-182 OLGU SUNUMU © Gülhane Askeri Týp Akademisi 2006 Azygos vein system abnormality: case report Necdet Kocabýyýk (*), Tunç Kutoðlu (**), Soner Albay (*), Bülent Yalçýn (*), Hasan Ozan (*) Summary Introduction Variations seen in the thoracic vein system are Abnormalities related to the azygos system are not rare (1). In a series related to the development of these veins. of 200 cases, Bergman et al. have reported the incidence of this anomaly During the dissection from the posterior medi- astinum of the 60-year-old male cadaver, it 26% (2). These abnormalities are generally explained by the embryolog- was observed that there was no complete ical development. Venous branching of the azygos vein varies (3). There accessory hemiazygos vein, and both posterior are two origins of the azygos and hemiazygos veins. By union of these intercostal veins and hemiazygos vein (above origins and regression of some parts, azygos system comes into its final T10 level) drained bilaterally to the azygos vein. Considering these types of variations is status (4). Different types of structures may occur when these veins important during imaging this region and surgi- develop. Abnormalities about azygos system and especially the variations cal operations. of the hemiazygos veins are not clearly described in the literature. In this Key words: Azygos vein, hemiazygos vein, superior vena cava, venous anomaly presentation absence of the accessory hemiazygos vein and possible causes of these types of variations are discussed in view of the embry- Özet ological development. Azigos ven sistem anomalisi: olgu sunumu Toraks ven sisteminde görülen varyasyonlar, embriyolojik olarak bu venlerin geliþimiyle ilgi- Case Report lidir. -

Analysis of Multiple Variations in Azygos Venous System Anatomy with Its Classification: a Cadaveric Study
ORIGINAL ARTICLE Eur. J. Anat. 23 (1): 9-15 (2019) Analysis of multiple variations in azygos venous system anatomy with its classification: A cadaveric study Apurba Patra1, Rajan K. Singla2, Harsimarjit Kaur2, Vishal Malhotra3 1Department of Anatomy, Dr Radhakrishnan, Government Medical College, Hamirpur (HP), India, 2Department of Anatomy, Government Medical College, Patiala, India, 3Department of SPM, Government Medical College, Patiala, India SUMMARY INTRODUCTION The azygos venous system varies greatly in The azygos system (gr. azygos – ‘unpaired’ or mode of its origin, course, number of vertical chan- ‘single’) which acts as a by-pass between the infe- nels, number of horizontal anastomoses and na- rior and superior vena caval systems (Bowsher, ture of termination. Anatomical knowledge of such 1954) is formed by veins which drain the posterior variations is of immense importance in radiological wall of the thorax and abdomen into the superior investigations and surgical intervention of posterior vena cava (SVC) (Tatar et al., 2008). It presents a mediastinum pathologies. The present study was tortuous appearance and usually lies anterior to undertaken on 30 adult embalmed cadavers aging the bodies of the thoracic vertebrae. The azygos between 40–65 years, to determine the anatomical system consists of three interconnected major variations of the azygos system and to classify veins, the azygos (AV), hemiazygos (HV) and ac- accordingly. The vertebral level and diameter of cessory hemiazygos veins (AHV) (Snell, 2004; the azygos, hemiazygos, accessory hemiazygos Drake et al., 2005). The AV begins on the posterior veins at their origin and terminations were also abdominal wall usually as a continuation of lumbar observed. -

The Azygos Vein System in the Rat
THE AZYGOS VEIN SYSTEM IN THE RAT MYRON H. HALPERN' Departments of Anatomy, University of Xichigan, Ann Arbor, and Hahnemann Medical College, Philadelphia THREE FIGURES The adult pattern of the azygos vein system of various mam- mals has received the attention of many early investigators (Eustachius, 1561; Bardeleben, 1848 ; Marshall, 1850; and Morrison, 1879). Since there has not almways been agreement among these workers in the patterns described, attempts were made by some of them to try to correlate the patterns on a de- velopmental basis (Barcleleben, 1848 ; Marshall, 1850; and Parker and Tozier, 1897). Tihey ( 'B), Kampmeier ( 'la), Sabin ( '14, '15), and Reagan ('19) each described the develop- ment of the azygos system of a different mammal. Although there was partial agreement on certain aspects of the embry- ology, it was not until recently that there has been general accord. Since one of the more significant contributions to the development of the azygos system in the rat (Strong, '36) has never appeared as a journal article and is procurable only as a thesis from the Indiana University Library, it will be included and related to the present description of the adult pattern. MATERIALS To investigate the constancy of pattern and to check fully the points of previous disagreement in the adult pattern, 57 rats were studied by the fluorescent-latex injection technique previously described by the author ( '52). Eleven additional 1 The author wishes to thank Dr. Russell T. Woodburne, Department of Anatomy, University of Michigan for his critical reading of this manuscript. a Portion of a dissertation submitted in partial fulfillment of the requirements for the degree of Doctor of Philosophy in the University of Michigan. -

What Is the History of the Term “Azygos Vein” in the Anatomical Terminology?
Surgical and Radiologic Anatomy (2019) 41:1155–1162 https://doi.org/10.1007/s00276-019-02238-3 REVIEW What is the history of the term “azygos vein” in the anatomical terminology? George K. Paraskevas1 · Konstantinos N. Koutsoufianiotis1 · Michail Patsikas2 · George Noussios1 Received: 5 December 2018 / Accepted: 2 April 2019 / Published online: 26 April 2019 © Springer-Verlag France SAS, part of Springer Nature 2019 Abstract The term “azygos vein” is in common use in modern anatomical and cardiovascular textbooks to describe the vein which ascends to the right side of the vertebral column in the region of the posterior mediastinum draining into the superior vena cava. “Azygos” in Greek means “without a pair”, explaining the lack of a similar vein on the left side of the vertebral column in the region of the thorax. The term “azygos” vein was utilized frstly by Galen and then was regenerated during Sylvius’ dissections and Vesalius’ anatomical research, where it received its fnal concept as an ofcial anatomical term. The purpose of this study is to highlight the origin of the term “azygos vein” to the best of our knowledge for the frst time and its evolu- tion from the era of Hippocrates to Realdo Colombo. Keywords Anatomy · “azygos vein” · “sine pari vena” · Terminology · Vesalius Introduction History of the origin of the term “azygos vein” The term “azygos vein” can be found in all modern ana- tomical textbooks. The term is used to describe a vein that Hippocrates (Fig. 1) did not make any mention with regard ascends on the right side of the vertebral column in the to the azygos vein. -

Double Inferior Vena Cava with Variant Hemiazygos Vein – a Case Report
IJAE Vol. 122, n. 2: 121-126, 2017 ITALIAN JOURNAL OF ANATOMY AND EMBRYOLOGY Research article - Human anatomy case report Double inferior vena cava with variant hemiazygos vein – a case report Sumathilatha Sakthi Velavan*, Bedia Castellanos, Sushama Rich, Robert Goldberg Department of Anatomy, Touro College of Osteopathic Medicine, Harlem, New York, NY – 10027 Abstract The duplication of the inferior vena cava is a rare variation resulting from an alteration in the embryogenesis of the cardinal venous system. Although there are various types of double infe- rior vena cava and is prevalent in 2-3% of the population, the continuation of left inferior vena cava as hemiazygos vein is a very unusual variant and hence this case is reported for its rar- ity and clinical significance. During dissection of an eighty-seven-year-old female cadaver, the presence of the double inferior vena cava was noted. A detailed dissection was done of the major veins of the abdomen and traced till their drainage into the thorax. The right and left inferior vena cava were connected by a venous bridge which coursed deep to the abdominal aorta. The right inferior vena cava followed its usual course and drained into the right atrium, while the left inferior vena cava entered the thoracic cavity as the hemiazygos vein and drained into the azygos vein. Anatomical knowledge of the rare variant prevents misdiagnosis and aids in the proper interpretation of radiological images. Also, awareness of this vascular anomaly guides the surgeons during retroperitoneal procedures when encountering intraoperative dif- ficulties. Key words Duplication, inferior vena cava, double IVC, azygos vein, hemiazygos vein. -

Anomalous Azygos Veins - Its Embryological Basis and Clinical Significance
International Journal of Research in Medical Sciences Shivanal U et al. Int J Res Med Sci. 2015 Sep;3(9):2323-2326 www.msjonline.org pISSN 2320-6071 | eISSN 2320-6012 DOI: http://dx.doi.org/10.18203/2320-6012.ijrms20150624 Research Article Anomalous azygos veins - its embryological basis and clinical significance Uma Shivanal1*, Geethanjali HT2 1Department of Anatomy, Mysore Medical College and Research Institute, Mysore, Karnataka, India 2Department of Anatomy, Mandya Institute of Medical Sciences, Mandya, Karnataka, India Received: 15 July 2015 Accepted: 10 August 2015 *Correspondence: Dr. Uma Shivanal, E-mail: [email protected] Copyright: © the author(s), publisher and licensee Medip Academy. This is an open-access article distributed under the terms of the Creative Commons Attribution Non-Commercial License, which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited. ABSTRACT Background: The anatomical knowledge of the variability of the azygos venous system is important for the surgical interventions of the posterior mediastinum and also during radiological investigations/diagnosis especially CT and MRI. The variant azygos venous system might be confused with thoracic aorta aneurysms, lymphadenopathy and tumours of posterior mediastinum. Methods: The present study was undertaken on 10 embalmed adult human cadavers irrespective of sex, used for undergraduate dissection from the Department of Anatomy, Mandya Institute of Medical Sciences, Mandya. In this present study, formation, course and termination pattern of azygos system of veins was observed in 10 dissected human cadavers. Out of which 2 cadavers showed different types of variations. Results: In the present study, normal azygos venous system was found in 8 specimens accounting for 80%. -

Superior and Posterior Mediastina Reading: 1. Gray's Anatomy For
Dr. Weyrich G07: Superior and Posterior Mediastina Reading: 1. Gray’s Anatomy for Students, chapter 3 Objectives: 1. Subdivisions of mediastinum 2. Structures in Superior mediastinum 3. Structures in Posterior mediastinum Clinical Correlate: 1. Aortic aneurysms Superior Mediastinum (pp.181-199) 27 Review of the Subdivisions of the Mediastinum Superior mediastinum Comprises area within superior thoracic aperture and transverse thoracic plane -Transverse thoracic plane – arbitrary line from the sternal angle anteriorly to the IV disk or T4 and T5 posteriorly Inferior mediastinum Extends from transverse thoracic plane to diaphragm; 3 subdivisions Anterior mediastinum – smallest subdivision of mediastinum -Lies between the body of sternum and transversus thoracis muscles anteriorly and the pericardium posteriorly -Continuous with superior mediastinum at the sternal angle and limited inferiorly by the diaphragm -Consists of sternopericardial ligaments, fat, lymphatic vessels, and branches of internal thoracic vessels. Contains inferior part of thymus in children Middle mediastinum – contains heart Posterior mediastinum Superior Mediastinum Thymus – lies posterior to manubrium and extends into the anterior mediastinum -Important in development of immune system through puberty -Replaced by adipose tissue in adult Arterial blood supply -Anterior intercostals and mediastinal branches of internal thoracic artery Venous blood supply -Veins drain into left brachiocephalic, internal thoracic, and thymic veins 28 Brachiocephalic Veins - Formed by the -

Anomalous Accessory Hemi-Azyos System with Persistent Cranial Segment of Posterior Cardinal Vein - a Case Report
Original article Anomalous accessory hemi-azyos system with persistent cranial segment of posterior cardinal vein - A case report Mahato, NK.* Department of Anatomy, Sri Aurobindo Institute of Medical Sciences – SAIMS, Indore-Ujjain Highway, Bhawrasala, Indore, Madhya Pradesh, India *E-mail: [email protected] Abstract Anomalies of the azygos system of vein are not uncommon. The case presented here exhibits persistent cranial part of the left posterior cardinal vein. This embryological variation gives rise to a well developed venous channel to the left side of the aortic arch and the thoracic aorta and drains the upper six left intercostals spaces. The superior intercostal vein on the left drains into this venous channel. The channel itself drains into the left brachio-cephalic vein after crossing the arch of aorta vertically on its left. Any communication between this anomalous channel and the azygos system on the right couldn’t be found. The inferior hemi-azygos system on the left is rudimentary. This persistent cephalic part of the posterior cardinal vein serves as a communication between the superior hemi-azygos vein to the brachio-cephalic vein on the left side. The existence of such an anomaly, the position of this vein and its proximity to the structures near the arch of aorta should be taken into consideration to avert radiological mis-diagnosis and surgical complications. Keywords: azygos system, posterior cardinal vein, hemi-azygos vein, brachio-cephalic vein, oblique vein of left atrium. 1 Introduction Anomalies of the azygos system of veins are not above give rise to the left superior intercostal vein in unison. -
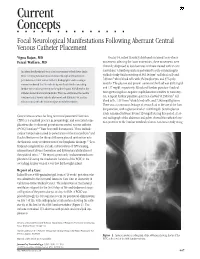
Current Concepts ⅢⅢⅢⅢⅢⅢⅢⅢⅢⅢⅢⅢⅢⅢ Focal Neurological Manifestations Following Aberrant Central Venous Catheter Placement
Current Concepts nnnnnnnnnnnnnn Focal Neurological Manifestations Following Aberrant Central Venous Catheter Placement Vigna Rajan, MD On day 14, infant B acutely developed sustained tonic-clonic Feizal Waffarn, MD movements affecting the lower extremities; these movements were clinically diagnosed as focal seizures and were treated with an anti- An infant developed focal tonic clonic movements of both lower limbs convulsant. A lumbar puncture performed to rule out meningitis 3 while receiving total parenteral nutrition through a left saphenous yielded cloudy fluid consisting of 34,114/mm red blood cells and 3 percutaneous central venous catheter. Radiographic studies using a 749/mm white blood cells with 3% lymphocytes and 97% poly- contrast confirmed that the catheter tip was located in the ascending morphs. The glucose and protein content of the fluid was 3943 mg/dl lumbar vein in close proximity to the epidural space. Withdrawal of the and 127 mg/dl, respectively. Blood and lumbar puncture fluid cul- catheter abated all clinical symptoms. This case emphasizes the need to tures grew coagulase-negative staphylococcus sensitive to vancomy- 3 confirm central venous catheter placement and illustrates yet another cin. A repeat lumbar puncture specimen showed 50,250/mm red 3 risk associated with the infusion of parenteral alimentation. blood cells, 1,515/mm white blood cells, and 7,348 mg/dl glucose. There was a continuous leakage of serous fluid at the site of the lum- bar puncture, with a glucose level of .800 mg/dl. Serum glucose levels remained between 80 and 120 mg/dl during this period. A lat- Central venous access for long-term total parenteral nutrition eral radiograph of the abdomen and pelvis showed the catheter loca- (TPN) is a standard practice in neonatology, and associated com- tion posterior to the lumbar vertebral column. -

Dilatation of the Azygos Vein Simulating a Mediastinal Tumour
Thorax: first published as 10.1136/thx.34.1.117 on 1 February 1979. Downloaded from Thorax, 1979, 34, 117-119 Dilatation of the azygos vein simulating a mediastinal tumour J A SIDDORN AND L WORSORNU From the Department of Medicine and Surgery, School of Medicine, University of Zambia, Lusaka, Zambia ABSTRACT A mediastinal tumour was simulated by gross dilatation of the azygos vein resulting from compression by enlarged hilar lymph nodes at the point where it crossed the right main bronchus to enter the superior vena cava. This cause of dilatation of the azygos vein has not previously been reported. Published reports are reviewed and attention drawn to the large proportion of patients (11/19) with this condition who had needless thoracotomies. The possibility of making a diagnosis by means of clinical observation and simple radiological techniques and thus avoiding surgical intervention is discussed. Dilatation of the azygos and hemiazygos veins had been present on the right side throughout his copyright. producing abnormal mediastinal or, less often, illness, and shortly before admission to hospital paraspinal shadows on chest radiographs is well he developed a constant dull gnawing pain deep to documented and has been reported in portal the supra-spinatus muscle. hypertension with collateral circulation (Leigh et He was thin, wasted, and appeared chronically al, 1954; Campbell and Baruch, 1960; Blendis et ill, but the only abnormal clinical finding was a al, 1968) and congenital and acquired abnor- right-sided pleural effusion. In particular there was http://thorax.bmj.com/ malities of both superior and inferior caval no engorgement of the neck veins, dependent systems (Shuford and Weens, 1958; Bogedain and oedema, varicosities of the legs, distension of Carpathios, 1962; Steinberg, 1962; Petersen, 1965; superficial collateral veins, palpable liver or spleen, Castellino et al, 1968; Floyd and Nelson, 1976) or evidence of hepatic disease.