HOW the COMMON BARN-OWL (Tyto Alba) HUNTS in DARKNESS by HEARING by H. Christian Floyd, Lexington How Do Owls Find and Catch
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Food-Niche Pattern of the Barn Owl (Tyto Alba) in Intensively Cultivated Agricultural Landscape˟
Ornis Hungarica 2018. 26(1): 27–40. DOI: 10.1515/orhu-2018-0002 Food-niche pattern of the Barn Owl (Tyto alba) in intensively cultivated agricultural landscape˟ Adrienn HORVÁTH, Anita MORVAI & Győző F. HORVÁTH* Received: February 28, 2018 – Revised: June 13, 2018 – Accepted: June 15, 2018 Horváth, A., Morvai, A. & Horváth, G. F. 2018. Food-niche pattern of the Barn Owl (Tyto alba) in intensively cultivated agricultural landscape. – Ornis Hungarica 26(1): 27–40. DOI: 10.1515/ orhu-2018-0002 ˟Presented at 1st Hungarian Owl Research Conference held in Pécs on 8th September 2017 Abstract This study investigated the dietary niche of the Barn Owl (Tyto alba) in an intensively farmed landscape, based on pellet samples from 12 nesting pairs containing 25 animal taxa and 1,994 prey items after the breeding sea- son in 2016. Based on land use categories of the buffer area around each nest, three landscape types (agricultural, mo- saic, urban) were considered, to analyse the diet composition and food-niche parameters. Niche breadth was calculat- ed at the local and landscape level. Small mammals were the most frequent in the diet than other prey in each of the landscape types. The Common Vole (Microtus arvalis), considered to be an important agricultural pest was the most numerous prey in all landscape groups. The trophic niche of Barn Owl varied between 0.69 – 0.86 at the local level, and the overall value of niche breadth was significantly higher in the urban than in the other two landscape types. Our results showed that the increase of Common Vole frequency lead to a decrease in niche breadth; significantly negative relationship was detected between these parameters. -

Tyto Longimembris) in Southern Taiwan
See discussions, stats, and author profiles for this publication at: https://www.researchgate.net/publication/251774474 Initial Investigation on the Diet of Eastern Grass Owl (Tyto longimembris) in Southern Taiwan ARTICLE in TAIWANIA · JANUARY 2007 CITATIONS READS 2 79 3 AUTHORS, INCLUDING: Hui-Yun Tseng 8 PUBLICATIONS 6 CITATIONS SEE PROFILE Available from: Hui-Yun Tseng Retrieved on: 14 January 2016 Taiwania, 52(1): 100-105, 2007 Initial Investigation on the Diet of Eastern Grass Owl (Tyto longimembris) in Southern Taiwan Wen-Loung Lin(1), Yin Wang(1) and Hui-Yun Tseng(2,3) (Manuscript received 4 October, 2006; accepted 26 December, 2006) ABSTRACT: This investigation, undertaken in the two regions of Nanshi and Yujing in Tainan County over the period of 2001 to 2003, included three nests belonging to the Eastern Grass Owl (Tyto longimembris). From these, we collected a total of 157 owl pellets. Analysis and examination of the pellets revealed 329 prey items. More in-depth investigation determined that 95.1% of the Eastern Grass Owl pellets collected consisted of mammal remains, while the remaining 4.9% were made up of bird remains. Of the various types of mammals consumed, rats made up the highest proportion, with a total of 285 rats, accounting for 86.6%. This was followed by 27 shrews and moles, accounting for 8.2%. Hares and birds were seldom caught and consumed. The findings suggested that rats are the main food source of the Eastern Grass Owl, with the Spinus Country-rat (Rattus losea) comprising the majority with 136 counted (41.3%), followed by the Formosan Mouse (Mus caroli) with 96 counted (29.2%). -

Tyto Alba (Barn Owl)
UWI The Online Guide to the Animals of Trinidad and Tobago Behaviour Tyto alba (Barn Owl) Family: Tytonidae (Barn Owls) Order: Strigiformes (Owls) Class: Aves (Birds) Fig. 1. Barn owl, Tyto alba. [http://aviary.owls.com/owls/barn-owl.html, downloaded 11 November 2012] TRAITS. There are approximately twenty different names used for this owl, each of which describes a particular feature of the owl or the owl’s appearance. For example, the monkey Faced owl is another name used for the barn owl because it sorts of resembles a monkey and rat owl because its main diet consists of rodents. Some other names include the white owl, ghost owl, screech owl, church owl, and night owl and so on (The Owl Pages, 2003). The scientific name “alba” means white and “tyto” means owl. The males are slightly smaller than the females. The female adult size ranges from 13.5-15.5 inches. The females also have a wing span of 43 inches and weigh 20 oz, whereas the male adult size ranges from 12.5-15 inches, a wing span of 42 UWI The Online Guide to the Animals of Trinidad and Tobago Behaviour inches and weighs 15.5 oz. These birds have distinct heads and white heart shaped faces which are outlined by a brownish colour (Harris, 2002). They do not have any ear tufts. The feathers at the upper surface of their body appear to be mostly golden with fine greyish streaks and dark spots. There are fewer dark spots on the breast and flanks on the underneath of the owl. -
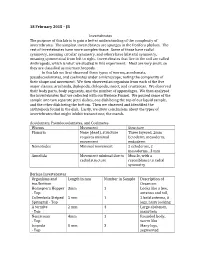
18 February 2015 – JB Invertebrates the Purpose of This Lab Is to Gain
18 February 2015 – JB Invertebrates The purpose of this lab is to gain a better understanding of the complexity of invertebrates. The simplest invertebrates are sponges in the Porifera phylum. The rest of invertebrates have more complex tissue. Some of those have radial symmetry, meaning circular symmetry, and others have bilateral symmetry, meaning symmetrical from left to right. Invertebrates that live in the soil are called Anthropods, which is what we studied in this experiment. Most are very small, so they are classified as microanthropods. In this lab we first observed three types of worms, acoelomats, pseudocoelomates, and coelmates under a microscope; noting the complexity of their shape and movement. We then observed an organism from each of the five major classes: arachnida, diplopoda, chilopoda, insect, and crustacean. We observed their body parts, body segments, and the number of appendages. We then analyzed the invertebrates that we collected with our Berlese Funnel. We poured some of the sample into two separate petri dishes, one dish being the top of our liquid sample, and the other dish being the bottom. Then we observed and identified the anthropods found in the dish. Lastly, we drew conclusions about the types of invertebrates that might inhibit transect one, the marsh. Acoelomats, Pseudocoelomates, and Coelmates Worms Movement Structure Planaria None (dead), structure Three layered, 2mm requires minimal Ectoderm, mesoderm, movement endoderm Nemotodes Minimal movement 2 ectoderms, 2 mesoderms , 3 mm Annelida Movement minimal due to Muscle, with a radial structure resemblance to radial symmetry Berlese Invertebrates Organisms and Length in mm Number in Sample Description of top/bottom Organism Homoptera Hopper 3mm 1 Looks like a bee, - Top antenna and tail, Collembola Striped 5 mm 1 2 bold antenna, 6 Springtail - Top legs, hairy looking A termite 2 mm 4 Large abdomen, - Top many legs Nematoma 4mm 1 Rounded body, - Top worm like Isopoda 8 mm 3 Many legs, - Top segmented The size range of the organisms observed was 2mm to 8mm. -

Strix Varia) from the Eastern United States T ⁎ Kevin D
Veterinary Parasitology: Regional Studies and Reports 16 (2019) 100281 Contents lists available at ScienceDirect Veterinary Parasitology: Regional Studies and Reports journal homepage: www.elsevier.com/locate/vprsr Original Article Trichomonosis due to Trichomonas gallinae infection in barn owls (Tyto alba) and barred owls (Strix varia) from the eastern United States T ⁎ Kevin D. Niedringhausa, ,1, Holly J. Burchfielda,2, Elizabeth J. Elsmoa,3, Christopher A. Clevelanda,b, Heather Fentona,4, Barbara C. Shocka,5, Charlie Muisec, Justin D. Browna,6, Brandon Munka,7, Angela Ellisd, Richard J. Halle, Michael J. Yabsleya,b a Southeastern Cooperative Wildlife Disease Study, 589 D.W. Brooks Drive, Wildlife Health Building, College of Veterinary Medicine, The University of Georgia, Athens, GA 30602, USA b Warnell School of Forestry and Natural Resources, 180 E Green Street, University of Georgia, Athens, GA 30602, USA c Georgia Bird Study Group, Barnesville, GA 30204, USA d Antech Diagnostics, 1111 Marcus Ave., Suite M28, Bldg 5B, Lake Success, NY 11042, USA e Odum School of Ecology and Department of Infectious Diseases, College of Veterinary Medicine, University of Georgia, Athens, GA 30602, USA ARTICLE INFO ABSTRACT Keywords: Trichomonosis is an important cause of mortality in multiple avian species; however, there have been relatively Owl few reports of this disease in owls. Two barn owls (Tyto alba) and four barred owls (Strix varia) submitted for Strix varia diagnostic examination had lesions consistent with trichomonosis including caseous necrosis and inflammation Tyto alba in the oropharynx. Microscopically, these lesions were often associated with trichomonads and molecular Trichomonosis testing, if obtainable, confirmed the presence of Trichomonas gallinae, the species most commonly associated Trichomonas gallinae with trichomonosis in birds. -

The Contribution of the Barn Owl (Tyto Alba) Feeding Ecology to Confirm Bat Species Occurrence in North Portugal
Hélia Marisa Vale-Gonçalves, Paulo Barros, Luís Braz, João Alexandre Cabral Barbastella 8 (1) Open Access ISSN: 1576-9720 SECEMU www.secemu.org The contribution of the Barn owl (Tyto alba) feeding ecology to confirm bat species occurrence in north Portugal Hélia Marisa Vale-Gonçalves1, Paulo Barros1, Luís Braz1 & João Alexandre Cabral1 1 Laboratory of Applied Ecology Centre for the Research and Technology of Agro-Environment and Biological Sciences (CITAB) University of Trás-os-Montes and Alto Douro (UTAD), Vila Real (Portugal). *Corresponding author e-mail: [email protected] DOI: http://dx.doi.org/10.14709/BarbJ.8.1.2015.05 © 2015 Published by SECEMU. Spanish title: Contribución de la ecología alimentaria de la lechuza común (Tyto alba) para confirmar la presencia de especies de quirópteros en el norte de Portugal Abstract: The Barn owl (Tyto alba) is an opportunistic species which feeds mainly on small mammals but also on birds, bats, reptiles, amphibians, insects and fishes. With regard to bats, several studies in Europe suggest that this group constitutes a small portion of the Barn owl diet representing less than 1% of its prey items. Through the analysis of 2,934 Barn owl pellets, collected between 2006 and 2014 in 27 sites/nests located in north Portugal, the remains of six bats belonging to five species were identified in a total of 9,103 prey items identified: the Western barbastelle (Barbastella barbastellus), the Grey long-eared bat (Plecotus austriacus), the Brown long-eared bat (Plecotus auritus), the European free-tailed bat (Tadarida teniotis) and the Common pipistrelle (Pipistrellus pipistrellus). -

A Revision of the Australian Owls (Strigidae and Tytonidae)
A REVISION OF THE AUSTRALIAN OWLS (STRIGIDAE AND TYTONIDAE) by G. F. MEES Rijksmuseum van Natuurlijke Historie, Leiden1) INTRODUCTION When in December 1960 the R.A.O.U. Checklist Committee was re- organised and the various tasks in hand were divided over its members, the owls were assigned to the author. While it was first thought that only the Boobook Owl, the systematics of which have been notoriously confused, would need thorough revision and that as regards the other species existing lists, for example Peters (1940), could be followed, it became soon apparent that it was impossible to make a satisfactory list without revision of all species. In this paper the four Australian species of Strigidae are fully revised, over their whole ranges, and the same has been done for Tyto tenebricosa. Of the other three Australian Tytonidae, however, only the Australian races have been considered: these species have a wide distribution (one of them virtually world-wide) and it was not expected that the very considerable amount of extra work needed to include extralimital races would be justified by results. Considerable attention has been paid to geographical distribution, and it appears that some species are much more restricted in distribution than has generally been assumed. A map of the distribution of each species is given; these maps are mainly based on material personally examined, and only when they extended the range as otherwise defined, have I made use of reliable field observations and material published but not seen by me. From the section on material examined it will be easy to trace the localities; where other information has been used, the reference follows the locality. -

Tasmanian Masked Owl)
The Minister included this species in the vulnerable category, effective from 19 August 2010 Advice to the Minister for Environment Protection, Heritage and the Arts from the Threatened Species Scientific Committee (the Committee) on Amendment to the list of Threatened Species under the Environment Protection and Biodiversity Conservation Act 1999 (EPBC Act) 1. Reason for Conservation Assessment by the Committee This advice follows assessment of new information provided through the Species Information Partnership with Tasmania on: Tyto novaehollandiae castanops [Tasmanian population] (Tasmanian Masked Owl) 2. Summary of Species Details Taxonomy Conventionally accepted as Tyto novaehollandiae castanops (Gould, 1837; Higgins, 1999; Christidis and Boles, 2008). There are three other subspecies of Tyto novaehollandiae which occur within Australia. Tyto novaehollandiae novaehollandiae occurs in southeast Queensland, eastern New South Wales, Victoria, southern South Australia and southern Western Australia. Tyto novaehollandiae kimberli is listed as vulnerable under the EPBC Act and occurs in northeast Queensland, the Northern Territory and northeast Western Australia. Tyto novaehollandiae melvillensis is listed as endangered under the EPBC Act and occurs on Melville Island and Bathurst Island (Higgins, 1999; DPIPWE, 2009). State Listing Status Listed as endangered under the Tasmanian Threatened Species Protection Act 1995. Description A large owl, weighing up to 1260 g, with a wingspan of up to 128 cm. Females are larger and heavier than males and considerably darker. The upperparts of this subspecies are dark brown to light chestnut in colour, with white speckling. The prominent facial disc is buff to chestnut coloured, with a darker margin, and chestnut coloured shading around the eyes. The legs are fully feathered and the feet are powerful with long talons (Higgins, 1999). -

(Tyto Alba) and Long-Eared Owl (Asio Otus) Mortality Along Motorways in Bourgogne-Champagne: Report and Suggestions
Barn Owl (Tyto alba) and Long-Eared Owl (Asio otus) Mortality Along Motorways in Bourgogne-Champagne: Report and Suggestions Hugues Baudvin1 The purpose of the study was to find where 305 km of motorways, from Dijon (Burgundy) and why two species of owls were killed by to Toul (Lorraine) and from Dijon to Saint- traffic along motorways. Three different factors Thibault (Champagne). Because of lack of have an important influence on the mortality of data, 46.1 km of road were not evaluated the two owl species: the biotops crossed by (discontinuous line figure 2). The distance motorways, the road elevation and the presence of small rodents, the Common Vole (Microtus arvalis) being most numerous. In order to limit the mortality caused by motorways, it is proposed to let the bordering vegetation grow naturally. Roadway-caused mortality of wildlife is a significant issue worldwide. Often, the impacts to wildlife occur along specific portions of roadways. Surveys along the roadways can identify these portions and identify the wildlife species being the most greatly impacted. Then, the management of roadside vegetation can help to reduce this negative impact. STUDY AREA AND METHODS The study area is located in the northeast of France, over the regions of Burgundy, Champagne, and Lorraine (fig. 1). It concerns Figure 2.—Owl mortality on motorways studied in northeastern France. studied is exactly 517.8 km (258.9 x 2 sides of the motorway). The study was conducted from November 1991 to December 1995. Dead animals were collected systematically along the motorway three times daily. The animals were placed in plastic bags and frozen for positive identification. -

Acoustic Location of Prey by Barn Owls {Tyto Alba) by Roger S
J. Exp. Biol. (1971), 54. S3S-573 535 With 5 plates and 5 text-figures Printed in Great Britain ACOUSTIC LOCATION OF PREY BY BARN OWLS {TYTO ALBA) BY ROGER S. PAYNE Rockefeller University and New York Zoological Society (Received 20 January 1970) INTRODUCTION The external ears of many species of owls are asymmetrical (Ridgeway, 1914; Stell- bogen, 1930). Although the asymmetry may involve the modification of different structures in different species (Stellbogen, 1930) the result is usually the same: one ear has its opening above the horizontal plane, the other below it. Though Pumphrey (1948), Southern (1955) and others have suggested that the asymmetrical ears of owls may be useful in hunting, prey location by hearing was first demonstrated by Payne & Drury (1958). Work prior to that time had suggested that, in the wild, light levels must often fall below that at which owls can see their prey, thus forcing them to rely upon some non-visual sense in hunting. Dice (1945) calculated the minimum intensi- ties of light required by the barn owl (Tyto alba), the long-eared owl (Asio otus), the barred owl (Strix varia), and the burrowing owl (Speotyto cunicularia) to see a dead mouse from a distance of 6 ft. His rough calculations of the reduction of light due to the absence of the moon, the shade of trees and shrubs, and various degrees of cloud cover led him to conclude that '.. .in the natural habitat of owls the intensity of illumination must often fall below the minimum at which the birds can see their prey'. -

Nesting Success of the Barn Owl (Tyto Alba) Using Nest Boxes in Southwest Missouri
BearWorks MSU Graduate Theses Summer 2017 Nesting Success Of The Barn Owl (Tyto Alba) Using Nest Boxes In Southwest Missouri Robert Grant Dickerson Missouri State University, [email protected] As with any intellectual project, the content and views expressed in this thesis may be considered objectionable by some readers. However, this student-scholar’s work has been judged to have academic value by the student’s thesis committee members trained in the discipline. The content and views expressed in this thesis are those of the student-scholar and are not endorsed by Missouri State University, its Graduate College, or its employees. Follow this and additional works at: https://bearworks.missouristate.edu/theses Part of the Behavior and Ethology Commons, Ornithology Commons, Population Biology Commons, and the Terrestrial and Aquatic Ecology Commons Recommended Citation Dickerson, Robert Grant, "Nesting Success Of The Barn Owl (Tyto Alba) Using Nest Boxes In Southwest Missouri" (2017). MSU Graduate Theses. 3190. https://bearworks.missouristate.edu/theses/3190 This article or document was made available through BearWorks, the institutional repository of Missouri State University. The work contained in it may be protected by copyright and require permission of the copyright holder for reuse or redistribution. For more information, please contact [email protected]. NESTING SUCCESS OF THE BARN OWL (TYTO ALBA) USING NEST BOXES IN SOUTHWEST MISSOURI A Masters Thesis Presented to The Graduate College of Missouri State University TEMPLATE In Partial Fulfillment Of the Requirements for the Degree Master of Science, Biology By Robert Grant Dickerson September 2017 Copyright 2017 by Robert Grant Dickerson ii NESTING SUCCESS OF THE BARN OWL (TYTO ALBA) USING NEST BOXES IN SOUTHWEST MISSOURI Biology Missouri State University, May 2017 Master of Science Robert Grant Dickerson ABSTRACT The Barn Owl (Tyto alba) is a medium-sized owl belonging to the family Tytonidae. -

Tyto Glaucops) Nest and First Record of Ashy-Faced Owl Nesting in Palmchat (Dulus Dominicus) Nest on Hispaniola
The Journal of Caribbean Ornithology RESEARCH NOTE Vol. 31:17–19. 2018 First description of Ashy-faced Owl (Tyto glaucops) nest and first record of Ashy-faced Owl nesting in Palmchat (Dulus dominicus) nest on Hispaniola Marta Curti Christine D. Hayes Thomas I. Hayes Misael Calcaño Silven Photo: Thomas I. Hayes The Journal of Caribbean Ornithology jco.birdscaribbean.org ISSN 1544-4953 RESEARCH NOTE Vol. 31:17–19. 2018 www.birdscaribbean.org First description of Ashy-faced Owl Tyto( glaucops) nest and first record of Ashy-faced Owl nesting in Palmchat Dulus( dominicus) nest on Hispaniola Marta Curti1,2, Christine D. Hayes1,3, Thomas I. Hayes1,4, and Misael Calcaño Silven1,5 Abstract Ashy-faced Owl (Tyto glaucops) is a Hispaniolan endemic. Despite the fact that it is considered locally common and widespread throughout much of its range, little is known about several aspects of this species’ biology, including its nesting habits. Its nest sites are described in general terms—e.g., tree cavities, caves, ledges—but no specific details, such as nest mea- surements, height above ground, type of substrate, or other information have been reported. We provide the first description of an Ashy-faced Owl nest and the first record of this species nesting in an active Palmchat (Dulus dominicus) nest. Keywords Ashy-faced Owl, Dulus dominicus, nest, nesting behavior, Palmchat, Tyto glaucops Resumen Primera descripción del nido de Lechuza Cara Ceniza (Tyto glaucops) y primer registro de nidificación de esta espe- cie en un nido de Cigua Palmera (Dulus dominicus) en La Española—La Lechuza Cara Ceniza (Tyto glaucops) es endémica a La Es- pañola.