Artikkel (1.496Mb)
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

The Anason Family in Rogaland County, Norway and Juneau County, Wisconsin Lawrence W
Andrews University Digital Commons @ Andrews University Faculty Publications Library Faculty January 2013 The Anason Family in Rogaland County, Norway and Juneau County, Wisconsin Lawrence W. Onsager Andrews University, [email protected] Follow this and additional works at: http://digitalcommons.andrews.edu/library-pubs Part of the United States History Commons Recommended Citation Onsager, Lawrence W., "The Anason Family in Rogaland County, Norway and Juneau County, Wisconsin" (2013). Faculty Publications. Paper 25. http://digitalcommons.andrews.edu/library-pubs/25 This Book is brought to you for free and open access by the Library Faculty at Digital Commons @ Andrews University. It has been accepted for inclusion in Faculty Publications by an authorized administrator of Digital Commons @ Andrews University. For more information, please contact [email protected]. THE ANASON FAMILY IN ROGALAND COUNTY, NORWAY AND JUNEAU COUNTY, WISCONSIN BY LAWRENCE W. ONSAGER THE LEMONWEIR VALLEY PRESS Berrien Springs, Michigan and Mauston, Wisconsin 2013 ANASON FAMILY INTRODUCTION The Anason family has its roots in Rogaland County, in western Norway. Western Norway is the area which had the greatest emigration to the United States. The County of Rogaland, formerly named Stavanger, lies at Norway’s southwestern tip, with the North Sea washing its fjords, beaches and islands. The name Rogaland means “the land of the Ryger,” an old Germanic tribe. The Ryger tribe is believed to have settled there 2,000 years ago. The meaning of the tribal name is uncertain. Rogaland was called Rygiafylke in the Viking age. The earliest known members of the Anason family came from a region of Rogaland that has since become part of Vest-Agder County. -

Hjelmeland 2021
Burmavegen 2021 Hjelmeland Nordbygda Velkomen til 2022 Kommunesenter / Municipal Centre Nordbygda Leite- Hjelmeland i Ryfylke Nesvik/Sand/Gullingen runden Gamle Hjelmelandsvågen Sauda/Røldal/Odda (Trolltunga) Verdas største Jærstol Haugesund/Bergen/Oslo Welcome to Hjelmeland, Bibliotek/informasjon/ Sæbø internet & turkart 1 Ombo/ in scenic Ryfylke in Fjord Norway Verdas største Jærstol Judaberg/ 25 Bygdamuseet Stavanger Våga-V Spinneriet Hjelmelandsvågen vegen 13 Sæbøvegen Judaberg/ P Stavanger Prestøyra P Hjelmen Puntsnes Sandetorjå r 8 9 e 11 s ta 4 3 g Hagalid/ Sandebukta Vågavegen a Hagalidvegen Sandbergvika 12 r 13 d 2 Skomakarnibbå 5 s Puntsnes 10 P 7 m a r k 6 a Vormedalen/ Haga- haugen Prestagarden Litle- Krofjellet Ritlandskrateret Vormedalsvegen Nasjonal turistveg Ryfylke Breidablikk hjelmen Sæbøhedlå 14 Hjelmen 15 Klungen TuntlandsvegenT 13 P Ramsbu Steinslandsvatnet Årdal/Tau/ Skule/Idrettsplass Hjelmen Sandsåsen rundt Liarneset Preikestolen Søre Puntsnes Røgelstad Røgelstadvegen KART: ELLEN JEPSON Stavanger Apal Sideri 1 Extra Hjelmeland 7 Kniv og Gaffel 10 SMAKEN av Ryfylke 13 Sæbøvegen 35, 4130 Hjelmeland Vågavegen 2, 4130 Hjelmeland Tlf 916 39 619 Vågavegen 44, 4130 Hjelmeland Tlf 454 32 941. www.apalsideri.no [email protected] Prisbelønna sider, eplemost Tlf 51 75 30 60. www.Coop.no/Extra Tlf 938 04 183. www.smakenavryfylke.no www.knivoggaffelas.no [email protected] Alt i daglegvarer – Catering – påsmurt/ Tango Hår og Terapi 2 post-i-butikk. Grocery Restaurant - Catering lunsj – selskapsmat. - Selskap. Sharing is Caring. 4130 Hjelmeland. Tlf 905 71 332 store – post office Pop up-kafé Hairdresser, beauty & personal care Hårsveisen 3 8 SPAR Hjelmeland 11 Den originale Jærstolen 14 c Sandetorjå, 4130 Hjelmeland Tlf 51 75 04 11. -

UNFC Case Studies from Finland/Estland, Sweden and Norway
United Nations ECE/ENERGY/GE.3/2020/10 Economic and Social Council Distr.: General 15 April 2020 Original: English only Economic Commission for Europe Committee on Sustainable Energy Expert Group on Resource Management Eleventh session Geneva, 20-24 April 2020 Item 11 of the provisional agenda Application of the United Nations Framework Classification for Resources (UNFC) in different geographical contexts. United Nations Framework Classification for Resources Case Studies from Finland/Estland, Sweden and Norway – Nordkalk limestone and Forsand sand and gravel mines Prepared by the Geological Survey of Norway, the Geological Survey of Sweden, Nordkalk, Forsand Sandkompani and Petronavit a.s. Summary This study demonstrates how the United Nations Framework Classification for Resources (UNFC) is being used to classify industrial mineral extraction projects for limestone in Estland and Sweden, and for a gravel and sand project in Norway. The report summarizes the main features of UNFC before describing the Nordkalk limestone projects. These projects demonstrate the usefulness of UNFC in categorizing environmental, social economic conditions and technical feasibility that may facilitate or hamper project development. The gravel and sand project demonstrates the use of UNFC in a case with few complications. This also shows an accounting method based on a standard that is common in national statistics as well as in project management. The gravel and sand case is also used to show how a UNFC inventory built with respect for the quality requirements embedded in the Committee for Mineral Reserves International Reporting Standards (CRIRSCO) may be translated into a CRIRSCO inventory. This allows for new UNFC accounts to be compared with existing CRIRSCO accounts and thus prevents a break in the records when going from a CRIRSCO standard to the UNFC standard ECE/ENERGY/GE.3/2020/10 Contents Page I. -

Kulturminneplan for Suldal
Kultur- minne- plan for Suldal Suldal kommune 2018 1 Kulturminneplan for Suldal Fagplan Vedtatt av kommunestyret 19.06. 2018 Digital versjon av planen er tilgjengeleg her: http://www.miljostatus-suldal.no/natur-og-kultur/ kulturminne-og-kulturlandskap/ Utarbeiding og layout: Naturinform SA, Sand Fotokrediteringar: Alle fotografi/illustrasjonar av Rune Roalkvam med følgjande unnatak (sidetal): Torbjørn Guggedal: 50 stort Tor Inge Havrevoll: 48 stort Alf Næsheim: 18 øvst Alf Odden: 20 to små. Bjarte Skipevåg: 33 (alle små bilde), 37 nedst, 38 øvst og nedst, 51 lite, 52 lite øvst, 56 stort. Suldal Sogelag: svartkvit bilde på s. 24, 31, 40, 43, 44, 45, 48, 49, 57, 59, 67, 70 Univ. oldsakssaml. Oslo: 18 nedst. Arkeologisk Museum Stavanger: 21 lite Gidske Vik: 35 stort Nasjonalgalleriet: 58 Statkart: 3, 60 Suldal kommune: 61 øvst. Illustrasjonar av publikasjonar m.v. vesentleg henta frå nettsider. Suldalsbeten 2 Innhald Føreord 4 Framtid med fotfeste – kulturminnepolitikken 6 1. Innleiing 8 1.1. Mål for kulturminneplanen 8 1.2. Plantype 8 1.3. Prosess 8 1.4. Planstatus 8 1.5. Kvifor ta vare på kulturminne? 9 1.6. Overordna føringar 10 1.7. Kva trugar kulturminna? 11 1.8. Aktørar, støtteordningar og virkemiddel 13 2. Kulturhistorisk oversyn 18 2.1. Fornminne 18 2.2. Bygningsmiljø, bygg og anlegg 26 2.3. Kulturlandskap 51 2.4. Gjenstandar 56 2.5. Immateriell kulturarv 62 3. Planverktøy 70 3.1. Mål for kulturminnevernet i Suldal 70 3.2. Strategiar 70 3.3. Retningsliner for bruk av kommunale midlar 72 3.4. Reglar for tildelte midlar 73 Vedlegg 1. Hus i SEFRAK kategori A 74 2. -

Rennesøy Finnøy Bokn Utsira Karmøy Tysvær Haugesund Vindafjord
227227 ValevValevåg Breiborg 336 239239 Ekkje HellandsbygHellandsbygd 236236 Hellandsbygda Utbjoa SaudaSauda 335 Etne Sauda Bratland 226226 Espeland VihovdaVihovda Åbødalen Brekke Sandvikdal Roaldkvam 240240 241241 Kastfosskrys 334 Egne Hjem UtbjoaUtbjoa Saunes 236 Skartland sør EtneEtne en TTråsavikikaaNærsonersone set 225225 Saua Førderde Gard Kvame Berge vest Ølen kirke Ølen Ørland Hytlingetong Gjerdevik ø EiodalenØlen skule den 332 SveioSveio 222222 Ølensvåg 235235 333 st Hamrabø Bråtveit Saudafjor 237237 224224 Øvrevre VatsVats SvandalSvandal MaldalMaldal 223223 Vindafjord SandeidSandeid Ulvevne Øvre Vats Skole Bjordalsveien Sandeid Fjellgardsvatnet Hylen Landa Løland Tengesdal 223 331 Knapphus Hordaland Østbø Hylen Blikrabygd 238238 Sandvik VanvikVanvik 328328 Vindafjord Hylsfjorden Skrunes SuldalseidSuldalseid Skjold 216216 218218 Sandeidfjorden 230230 Suldal NordreNordre VikseVikse Ørnes 228228 RopeidRopeid Isvik SkjoldSkjold Hustoft VikedalVikedal 217217 Slettafjellet Nesheim Vikedal Helganes 228 330330 Vikse kryss VestreVestre Skjoldafjo Eskedalen 221221 Suldalseid SuldalsosenSuldalsosen Haugesund Stølekrossen Førland Åmsoskrysset Åmsosen IlsvIlsvåg Stole Ølmedal Kvaløy Ropeid Skipavåg Sand Suldalsosen rden 230 Byheiene 229229 Roopeid 327 327327 Stakkestad SkipevSkipevåg kryss SandSand Kariås Røvær kai Nesheim Eikanes kryss Lindum Årek 329329 camping 602 VasshusVasshus Røvær 200200 220220 Suldalsl HaugesundHaugesund ågen Haraldshaugen 205205 Kvitanes Grindefjorden NedreNedre VatsVats Vindafjorden Kvamen Haugesund Gard skole -

Annual Report Research Activity 2019
Annual Report Research Activity 2019 Division of Clinical Neuroscience University of Oslo and Oslo University Hospital 0 Contents Oslo University Hospital and the University of Oslo .................................................................................... 4 From Division Director Eva Bjørstad ........................................................................................................... 4 Division of Clinical Neuroscience (NVR) Organizational Chart ..................................................................... 5 Department of Physical Medicine and Rehabilitation Rehabilitation after trauma....................................................................................................................... 6 Group Leader: Nada Andelic Painful musculoskeletal disorders .............................................................................................................. 9 Group Leader: Cecilie Røe Department of Refractory Epilepsy - National Centre for Epilepsy Complex epilepsy .................................................................................................................................... 11 Group Leader: Morten Lossius Department of Neurosurgery Neurovascular-Hydrocephalus Research Group ..................................................................................... 16 Group Leader: Per Kristian Eide Oslo Neurosurgical Outcome Study Group (ONOSG) ................................................................................. 19 Group Leaders: Eirik Helseth and Torstein -

Larson (1886-1957); from the Bakken Subfarm, Guggedal Main Farm
THE NORWEGIAN ANCESTRY OF JOHANNES (JOHN) LARSON (1886-1957); FROM THE BAKKEN SUBFARM, GUGGEDAL MAIN FARM IN ROGALAND COUNTY, NORWAY TO THE SULDAL NORWEGIAN SETTLEMENT IN JUNEAU COUNTY, WISCONSIN John Larson (#275) and Lars Benson (see #99) Crossing of Suldal and Johnson Roads, Lindina Township, Juneau County, Wisconsin BY LAWRENCE W. ONSAGER THE LEMONWEIR VALLEY PRESS Berrien Springs, Michigan and Mauston, Wisconsin 2018 1 COPYRIGHT (C) 2018 by Lawrence W. Onsager All rights reserved including the right of reproduction in whole or in part in any form, including electronic or mechanical means, information storage and retrieval systems, without permission in writing from the author. Manufactured in the United States of America ------------------------------------------------------------------------------------------------------ Cataloging in Publication Data Onsager, Lawrence William, 1944- The Norwegian Ancestry of Johannes (John) Larson (1886-1957) From the Bakken Subfarm, Guggedal Main Farm in Rogaland County, Norway to the Suldal Norwegian Settlement in Juneau County, Wisconsin, Mauston, Wisconsin and Berrien Springs, Michigan: The Lemonweir Valley Press, 2018. 1. Juneau County, Wisconsin 2. Larson Family 3. Suldal Parish, Rogaland County, Norway 4. Onsager Family 5. Ormson Family 6. Juneau County, Wisconsin – Norwegians 7. Gran Parish, Oppland County, Norwa I. Title Series: Suldal Norwegian-American Settlement, Juneau County, Wisconsin Tradition claims that the Lemonweir River was named for a dream. Prior to the War of 1812, an Indian runner was dispatched with a war belt of wampum with a request for the Dakotas and Chippewas to meet at the big bend of the Wisconsin River (Portage). While camped on the banks of the Lemonweir, the runner dreamed that he had lost his belt of wampum at his last sleeping place. -

Det-Norske-Myrselskap-1949
MEDDELELSER FRA DET NORSl(E MYRSELSl(AP Nr. 6 Desember 1949 47. årgang Redigert av Aasulv Løddesøl. MYRENE I KYSTHERREDENE I NORD-ROGALAND. Av konsulent Ose. Hovde. Nord-Rogaland er vanlig benevnelse for den del av Rogala,nd fylke som ligger nord for Boknafjorden. De typiske kystherreder i Nord-Rogaland - regnet nordfra -- er følgende 8: Skåre, Torvastad, Avaldsnes, Utsira, Åkra, Stangaland, Skudenes og Bokn. Innen dette kystområde ligger også 3 bykommuner, nemlig Haugesund, Kopervik og Skudeneshavn. Nærmere geografisk bestemt ligger disse herreder mellom 59°08' og 59°31' nordlig bredde og mellom 5~10' og 6° vestlig lengde (Oslo meridian). Herredenes totalareal er 407,96 km2 og land• arealet 392,02 km2• Av hele Rogaland fylke utgjør dette område 4,50 %. Det meste av Skåre og Avaldsnes herreder med Haugesund by og en mindre del av Torvastad ligger på fastland (Haugalandet), men for øvrig ligger de andre herreder på øyer. Den største av disse er Karmøya. Her ligger de tre herreder Skudenes, Akra og Stanga• land i sin helhet og dessuten det meste av Torvastad og en del av Avaldsnes. Her er dessuten de to bykommuner Kopervik og Skude·• neshavn. Karmøya er således Norges folkerikeste og tettest bebodde øy. Bokn herred ligger på flere øyer øst for Karmøya, og Utsira her• red ligger langt ute i havet (16 km) rett vest av Karmøya, Kommunikasjonene innen området er stort sett gode, men den svære trafikk over Karmsundet ·gjør krav om bru eller bedre ferje• forbindelse berettiget. Ruten på Karmøya trafikeres med busser av Norges Statsbaner. F j e 11 grunnen innen området består stort sett av 3 temmelig skarpt atskilte områder. -
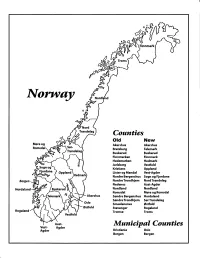
Norway Maps.Pdf
Finnmark lVorwny Trondelag Counties old New Akershus Akershus Bratsberg Telemark Buskerud Buskerud Finnmarken Finnmark Hedemarken Hedmark Jarlsberg Vestfold Kristians Oppland Oppland Lister og Mandal Vest-Agder Nordre Bergenshus Sogn og Fjordane NordreTrondhjem NordTrondelag Nedenes Aust-Agder Nordland Nordland Romsdal Mgre og Romsdal Akershus Sgndre Bergenshus Hordaland SsndreTrondhjem SorTrondelag Oslo Smaalenenes Ostfold Ostfold Stavanger Rogaland Rogaland Tromso Troms Vestfold Aust- Municipal Counties Vest- Agder Agder Kristiania Oslo Bergen Bergen A Feiring ((r Hurdal /\Langset /, \ Alc,ersltus Eidsvoll og Oslo Bjorke \ \\ r- -// Nannestad Heni ,Gi'erdrum Lilliestrom {", {udenes\ ,/\ Aurpkog )Y' ,\ I :' 'lv- '/t:ri \r*r/ t *) I ,I odfltisard l,t Enebakk Nordbv { Frog ) L-[--h il 6- As xrarctaa bak I { ':-\ I Vestby Hvitsten 'ca{a", 'l 4 ,- Holen :\saner Aust-Agder Valle 6rrl-1\ r--- Hylestad l- Austad 7/ Sandes - ,t'r ,'-' aa Gjovdal -.\. '\.-- ! Tovdal ,V-u-/ Vegarshei I *r""i'9^ _t Amli Risor -Ytre ,/ Ssndel Holt vtdestran \ -'ar^/Froland lveland ffi Bergen E- o;l'.t r 'aa*rrra- I t T ]***,,.\ I BYFJORDEN srl ffitt\ --- I 9r Mulen €'r A I t \ t Krohnengen Nordnest Fjellet \ XfC KORSKIRKEN t Nostet "r. I igvono i Leitet I Dokken DOMKIRKEN Dar;sird\ W \ - cyu8npris Lappen LAKSEVAG 'I Uran ,t' \ r-r -,4egry,*T-* \ ilJ]' *.,, Legdene ,rrf\t llruoAs \ o Kirstianborg ,'t? FYLLINGSDALEN {lil};h;h';ltft t)\l/ I t ,a o ff ui Mannasverkl , I t I t /_l-, Fjosanger I ,r-tJ 1r,7" N.fl.nd I r\a ,, , i, I, ,- Buslr,rrud I I N-(f i t\torbo \) l,/ Nes l-t' I J Viker -- l^ -- ---{a - tc')rt"- i Vtre Adal -o-r Uvdal ) Hgnefoss Y':TTS Tryistr-and Sigdal Veggli oJ Rollag ,y Lvnqdal J .--l/Tranbv *\, Frogn6r.tr Flesberg ; \. -

ROGALAND 23. JULY – 4. AUGUST 2014 Sunday the 27Th We Traveled
ROGALAND 23. JULY – 4. AUGUST 2014 Sunday the 27th we traveled to Karmøy. We drove Here we have parked. through the new tunnel in the triangular connection to Skudesneshavn. We stopped at Skudenes Camping. We parked at this place. The following day, Sunday the 28th, we took a trip down to Skudesneshavn. Those who live here can park their boat right outside Here we are at Kanalen. their door. View across the strait to Vaholmen. Terraces with moored boats. View south towards the bridge that goes over to Skudenes Mekaniske Verksted was built in 1916. Here Steiningsholmen. were the Skude engines produced. I the end of the house is written the name on the wall. Here we come to the square located down by the harbor. Skudeneshavn is a town and a former municipality in Karmøy municipality in Rogaland. Skudesneshavn had 3,327 inhabitants as of 1 January 2013, and is located on Skudeneset on the southern tip of the island Karmøy. Skudesneshavn grew up in the late Middle Ages and the urban society grew rapidly during the herring fishery in the 1800s.Today, the city has modern shipbuilding industry and one of the largest offshore shipping. Solstad Offshore has one of the world's most advanced offshore fleets in service throughout the world. The well-preserved wooden buildings along the harbor, Søragadå, has become one of the region's most visited tourist destination. The flounder fisherman The lobster fisherman The statues in Nordmand valley, Denmark In Nordmandsdalen in Fredensborg Palace Gardens stand 60 statues from various parts of Norway. -

Administrative and Statistical Areas English Version – SOSI Standard 4.0
Administrative and statistical areas English version – SOSI standard 4.0 Administrative and statistical areas Norwegian Mapping Authority [email protected] Norwegian Mapping Authority June 2009 Page 1 of 191 Administrative and statistical areas English version – SOSI standard 4.0 1 Applications schema ......................................................................................................................7 1.1 Administrative units subclassification ....................................................................................7 1.1 Description ...................................................................................................................... 14 1.1.1 CityDistrict ................................................................................................................ 14 1.1.2 CityDistrictBoundary ................................................................................................ 14 1.1.3 SubArea ................................................................................................................... 14 1.1.4 BasicDistrictUnit ....................................................................................................... 15 1.1.5 SchoolDistrict ........................................................................................................... 16 1.1.6 <<DataType>> SchoolDistrictId ............................................................................... 17 1.1.7 SchoolDistrictBoundary ........................................................................................... -

Forslag Til Forvaltningsplan for Deler Av Skudeneshavn
FORSLAG TIL FORVALTNINGSPLAN FOR SKUDENESHAVN FORSLAG TIL FORVALTNINGSPLAN FOR SKUDENESHAVN 1 Forvaltningsplanen er utarbeidet av Riksantikvaren 01.03.2015–05.11.2015 Prosjektleder: Ane Steingildra Alvestad (Karmøy kommune, innleid av Riksantikvaren i prosjektperioden) Tekst: Ane Steingildra Alvestad i samarbeid med arbeidsgruppa for Skudeneshavn kulturmiljø I tillegg til prosjektleder har arbeidsgruppa bestått av: Ingrid Djupedal (Riksantikvaren) Monica Rusten (Riksantikvaren) Cathrine Skredderstuen Rolland (Riksantikvaren) Elisabeth Nordling (Riksantikvaren) Eline Ova Sveen (Riksantikvaren) May Britt Håbjørg (Riksantikvaren) Jorun Aresvik Hals (Riksantikvaren) Lars Tveit (Karmøy kommune) Signe Oddbjørg Raen (Rogaland fylkeskommune) Foto: Ane Steingildra Alvestad Grafisk design: Ginungagap Trykk: Haugesund Bok & Offset Forslag til forvaltningsplan Skudeneshavn 2 FORSLAG TIL FORVALTNINGSPLAN FOR SKUDENESHAVN Innholdsfortegnelse Forord ................................................................................................................................................................................................ 9 DEL 1 INNLEDNING ..................................................................................................................................................................... 11 1 Forvaltningsplanens formål og innhold .................................................................................................. 13 2 Skudeneshavn – kort om verneverdier...................................................................................................