Swiftlet Behaviour Responses to Predators in Proximity to Their Nests
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

NSW Vagrant Bird Review
an atlas of the birds of new south wales and the australian capital territory Vagrant Species Ian A.W. McAllan & David J. James The species listed here are those that have been found on very few occasions (usually less than 20 times) in NSW and the ACT, and are not known to have bred here. Species that have been recorded breeding in NSW are included in the Species Accounts sections of the three volumes, even if they have been recorded in the Atlas area less than 20 times. In determining the number of records of a species, when several birds are recorded in a short period together, or whether alive or dead, these are here referred to as a ‘set’ of records. The cut-off date for vagrant records and reports is 31 December 2019. As with the rest of the Atlas, the area covered in this account includes marine waters east from the NSW coast to 160°E. This is approximately 865 km east of the coast at its widest extent in the south of the State. The New South Wales-Queensland border lies at about 28°08’S at the coast, following the centre of Border Street through Coolangatta and Tweed Heads to Point Danger (Anon. 2001a). This means that the Britannia Seamounts, where many rare seabirds have been recorded on extended pelagic trips from Southport, Queensland, are east of the NSW coast and therefore in NSW and the Atlas area. Conversely, the lookout at Point Danger is to the north of the actual Point and in Queensland but looks over both NSW and Queensland marine waters. -

Compile and Review Invasive Alien Species Information for the Cook Islands Report for the National Environment Service
Compile and Review Invasive Alien Species Information for the Cook Islands Report for the National Environment Service Coordinator of the Invasive Alien Species (IAS) in the Cook Islands Project Prepared by Shyama Pagad Invasive Species Specialist Group (ISSG) Pacific Regional Office January 2014 1 | Page Contents Compile and Review Invasive Alien Species Information for the Cook Islands ........................... 1 Glossary and Definitions .......................................................................................................................... 4 Introduction ............................................................................................................................................ 5 SECTION 1 ............................................................................................................................................... 8 Alien and Invasive Species in the Cook Islands ..................................................................................... 8 SECTION 2 ............................................................................................................................................. 11 Pathways of introduction and spread ................................................................................................ 11 SECTION 3 ............................................................................................................................................. 14 Cook Islands and its biodiversity ....................................................................................................... -

Echolocation in Oilbirds and Swiftlets
REVIEW ARTICLE published: 28 May 2013 doi: 10.3389/fphys.2013.00123 Echolocation in Oilbirds and swiftlets Signe Brinkløv 1*, M. Brock Fenton 1 and John M. Ratcliffe 2 1 Department of Biology, Western University, London, ON, Canada 2 Institute of Biology, University of Southern Denmark, Odense, Denmark Edited by: The discovery of ultrasonic bat echolocation prompted a wide search for other animal Mariana L. Melcón, Fundación biosonar systems, which yielded, among few others, two avian groups. One, the South Cethus, Argentina American Oilbird (Steatornis caripensis: Caprimulgiformes), is nocturnal and eats fruit. The Reviewed by: other is a selection of diurnal, insect-eating swiftlets (species in the genera Aerodramus Noritaka Ichinohe, National Institute of Neuroscience, Japan and Collocalia:Apodidae)fromacrosstheIndo-Pacific.Birdecholocationisrestrictedto Lore Thaler, Durham University, UK lower frequencies audible to humans, implying a system of poorer resolution than the *Correspondence: ultrasonic (>20 kHz) biosonar of most bats and toothed whales. As such, bird echolocation Signe Brinkløv, Department of has been labeled crude or rudimentary. Yet, echolocation is found in at least 16 extant Biology, Western University, bird species and has evolved several times in avian lineages. Birds use their syringes to 1151 Richmond Street, London, ON N6A 3K7, Canada. produce broadband click-type biosonar signals that allow them to nest in dark caves and e-mail: [email protected] tunnels, probably with less predation pressure. There are ongoing discrepancies about several details of bird echolocation, from signal design to the question about whether echolocation is used during foraging. It remains to be seen if bird echolocation is as sophisticated as that of tongue-clicking rousette bats. -

The Avifauna of Lambusango Forest Reserve, Buton Island, South-East Sulawesi, with Additional Sightings from Southern Buton
FORKTAIL 28 (2012): 107–112 The avifauna of Lambusango Forest Reserve, Buton Island, south-east Sulawesi, with additional sightings from southern Buton T. E. MARTIN, D. J. KELLY, N. T. KEOGH, D. HERIYADI, H. A. SINGER & G. A. BLACKBURN Lambusango Forest Reserve occupies a large area of south-central Buton, the largest attendant island of Sulawesi, Indonesia. Buton is located off Sulawesi’s south-eastern peninsula and remains poorly known ornithologically. Bird surveys were undertaken in the reserve over eight eight-week long research seasons between June and August in 1999, 2001–2003, 2005, and 2008–2010. Variable radius circular- plot point counts were the primary census method, conducted as part of a long-term biodiversity monitoring programme in the reserve, although data were also collected from 840 mist-netting hours and approximately 2,560 hours of observational data. In total, 79 species were detected in the reserve, including 37 regional endemics (46.8% of the total avifaunal community) and four species considered by the IUCN to be globally threatened or Near Threatened. Additionally, a further 60 species (including two more Near Threatened species) were recorded in various habitats around southern Buton that were not detected in Lambusango Reserve, giving a total of 139 species records for the island. We believe that 51 of these species represent previously unpublished records for Buton. We present here a full account of our records from Lambusango Reserve and southern Buton, with additional details provided for threatened and Near Threatened species and new records of endemics. INTRODUCTION Lambusango Forest Reserve (5°10’–5°24’S 122°43’–123°07’E) is a 65,000 ha expanse of uninhabited tropical monsoon forest, Buton (formerly referred to as Butung) is the largest of Sulawesi’s encompassing much of south-central Buton. -

Extinct and Extirpated Birds from Aitutaki and Atiu, Southern Cook Islands 1
Pacific Science (1991), vol. 45, no. 4: 325-347 © 1991 by University of Hawaii Press. All rights reserved Extinct and Extirpated Birds from Aitutaki and Atiu, Southern Cook Islands 1 DAVID W. STEADMAN 2 ABSTRACT: Six archaeological sites up to 1000 years old on Aitutaki, Cook Islands, have yielded bones of 15 species of birds, five of which no longer occur on the island: Pterodroma rostrata (Tahiti Petrel); Sula sula (Red-footed Booby); Dendrocygna, undescribed sp. (a large, extinct whistling duck); Porzana tabuensis (Sooty Crake); and Vini kuhlii (Rimatara Lorikeet). Of these, only S. sula and P. tabuensis survive anywhere in the Cook Islands today . The nearest record of any speciesofDendrocygna is in Fiji. Aside from the aquatic speciesEgretta sacra (Pacific Reef-Heron) and Anas superciliosa (Gray Duck), the only native, resi dent land bird on Aitutaki today is the Society Islands Lorikeet (Vini peru viana), which may have been introduced from Tahiti. Residents of Aitutaki note that Ducula pacifica (Pacific Pigeon) and Ptilinopus rarotongensis (Cook Islands Fruit-Dove) also occurred there until the 1940sor 1950s.There is no indigenous forest on Aitutaki today. The bones from Aitutaki also include the island's first record of the fruit bat Pteropus tonganus . Limestone caves on the island of Atiu yielded the undated bones of six species of birds, three of which no longer exist there or anywhere else in the Cook Islands: Gallicolumba erythroptera (Society Islands Ground-Dove), Ducula aurorae (Society Islands Pigeon), and Vini kuhlii. Each of these species has been recovered from prehistoric sites on Mangaia as well. The limestone terrain of Atiu is mostly covered with native forest that supports populations of Ducula pacifica, Ptilinopus rarotongensis, Collocalia sawtelli (Atiu Swiftlet), and Halcyon tuta (Chattering Kingfisher). -

Tropical & Far North Queensland 9 Day Birding Tour
Bellbird Tours Pty Ltd PO Box 2008, BERRI SA 5343 AUSTRALIA Ph. 1800-BIRDING Ph. +61409 763172 www.bellbirdtours.com [email protected] Tropical & Far North Queensland 9 day birding tour Tropical Far North Queensland is without doubt one of known for such as Magnificent Riflebird, Frill-necked Monarch, Australia‟s top birding destinations. A variety of tropical Monarch, Blue-faced Parrot-finch, Lovely Fairy-wren, White- habitats including rainforests, palm-fringed beaches, White-streaked Honeyeater, Papuan Frogmouth, nesting Red mangrove-lined mudflats, savannahs and cool mountain Red Goshawks, Palm Cockatoo, Eclectus Parrot, Yellow-bellied ranges result in a bird diversity unparalleled elsewhere in bellied Kingfisher and many more, including of course the rare the country. rare Southern Cassowary. We‟ll have rare access to Golden- Commencing in Cairns, we‟ll bird the local area and head shouldered Parrot nesting sites; explore Lakefield National to well-known Kingfisher Park on the Atherton Tablelands, Park; we‟ll go night spotting for owls and mammals‟; and we‟ll then traverse the unique Cape York Peninsula. During this we‟ll spend two days in Iron Range National Park, one of tour we aim to find all Tropical Far North Queensland Australia‟s most important ecosystems. We‟ll also look for specialties. Expect over 200 species including all the mammals such as Bandicoot, Sugar Glider, Northern Striped endemics and other important birds this area is well- Striped Possum, Red-necked Wallaby and Tree Kangaroo. Read on below for the full -

Palau Bird Survey Report 2020
Abundance of Birds in Palau based on Surveys in 2005 Final Report, November 2020 Eric A. VanderWerf1 and Erika Dittmar1 1 Pacific Rim Conservation, 3038 Oahu Avenue, Honolulu, Hawaii 96822 Prepared for the Belau National Museum, Box 666, Koror Palau 96940 Endemic birds of Palau, from top left: White-breasted Woodswallow, Palau Fantail, Palau Fruit- dove, Rusty-capped Kingfisher. Photos by Eric VanderWerf. 1 TABLE OF CONTENTS ACKNOWLEDGMENTS .............................................................................................................. 3 EXECUTIVE SUMMARY ............................................................................................................ 4 INTRODUCTION .......................................................................................................................... 5 METHODS ..................................................................................................................................... 6 Description of Study Area and Transect Locations ............................................................ 6 Data Collection ................................................................................................................... 7 Data Analysis ...................................................................................................................... 7 Limitations of the Survey.................................................................................................... 9 RESULTS .................................................................................................................................... -

Cooktown and District Bird Species
Cooktown and District Bird Species Southern Cassowary L White-necked Heron U Pacific Golden Plover C,M Pheasant Coucal C Yellow-spotted Honeyeater C Rufous Fantail U Emu R Eastern Great Egret C Grey Plover R,M Eastern Koel U,M,h Graceful Honeyeater U Grey Fantail U Australian Brush-turkey C Intermediate Egret U Red-capped Plover C Channel-billed Cuckoo U,M,h Bridled Honeyeater L,E Northern Fantail U Orange-footed Scrubfowl C Great-billed Heron L,R Lesser Sand Plover U,M Horsfield's Bronze-Cuckoo U,h Yellow-faced Honeyeater U Willie Wagtail U Brown Quail R Cattle Egret C Greater Sand Plover C,M Shining Bronze-Cuckoo U,h Varied Honeyeater L Torresian Crow C Indian Peafowl L,R Striated Heron U Black-fronted Dotterel U Little Bronze-Cuckoo C,h Yellow Honeyeater C Leaden Flycatcher C Magpie Goose U Pied Heron R,M Red-kneed Dotterel R Pallid Cuckoo U Brown-backed Honeyeater C Satin Flycatcher R,M Plumed Whistling-Duck R White-faced Heron C Masked Lapwing C Chestnut-breasted Cuckoo U Bar-breasted Honeyeater R Shining Flycatcher C Wandering Whistling-Duck U Little Egret C Comb-crested Jacana C Fan-tailed Cuckoo U Dusky Honeyeater C Restless Flycatcher R Radjah Shelduck U Eastern Reef Egret C Latham's Snipe U,M Brush Cuckoo C,h Scarlet Honeyeater U White-eared Monarch U Green Pygmy-goose C Nankeen Night-heron U Black-tailed Godwit R,M Oriental Cuckoo R,M Banded Honeyeater U Black-faced Monarch U Grey Teal U Glossy Ibis R Bar-tailed Godwit C,M Barking Owl U,h Brown Honeyeater R Black-winged Monarch L,M Pacific Black Duck C Australian White Ibis -

Volume 33 Number 1 March 2009
57772_No1_Pg1_6.qxd 18/2/09 9:34 AM Page 1 CORELLA Journal of the Australian Bird Study Association Volume 33 Number 1 March 2009 Corella, 2009, 33(1): 1-6 THE BREEDING BIOLOGY OF THE WHITE-RUMPED SWIFTLET Aerodramus spodiopygius spodiopygius IN SAMOA MICHAEL K. TARBURTON School of Science and Technology, Pacific Adventist University, PMB, Boroko, Papua New Guinea. Present address: 3 Freda Court, Blackburn South, Victoria 3130. Australia. E-mail: [email protected] Received: 2 August 2004 In the lava-tube caves of Samoa the White-rumped Swiftlet Aerodramus s. spodiopygius has a protracted breeding season, laying eggs every month of the year. It builds nests of vegetation and saliva that average 44.7 × 43.7 × 11.8 millimetres in size, 5.1 metres from the cave floor and 5.2 metres from their nearest neighbour. These nests took 19 days to three months to build and usually contained a single egg weighing 1.6 grams and 18.2 × 12.3 millimetres in size. The nests lasted an average of 9.4 months and were used to raise up to three broods. The eggs took 25 or 26 days to hatch and the nestlings fledged in 47–57 days. Nesting success was 72 per cent in the Tafatafa colony and 45 per cent in the Falemauga Small Cave colony. The 72 per cent success rate is the highest for any swiftlet studied to date. The 45 per cent rate was probably a result of a combination of low placement of some nests, infertility, predation by rats or disturbance by observers. -

Santo Edited by Philippe Bouchet, Hervé Le Guyader, Olivier Pascal the Natural History of N Aturels
Santo edited by Philippe Bouchet, Hervé Le Guyader, Olivier Pascal The Natural History of ATURELS N PUBLICATIONS SCIENTIFIQUES DU MUSÉUM IRD ÉDITIONS PATRIMOINES PRO-NATURA INTERNATIONAL The Santo 2006 expedition was organized, with the support, among others, of and Représentation de l'Union Européenne au Vanuatu Santo edited by Philippe Bouchet, Hervé Le Guyader & Olivier Pascal The Natural History of Foreword by Edward Natapei, Prime Minister of Vanuatu ...................................... 10 Introduction. The Natural History of Santo: An Attempt to Bridge the Gap between Academic Research and Consevation and Education .......................... 11 Vanuatu in the South Pacific ............................................................................................................. 13 Benoît Antheaume EspiritU santO in space AND time ........................................................................................... 17 coordinated by Bruno Corbara The Late Quaternaty Reefs ............................................................................................................................. 19 Guy Cabioch & Frederick W. Taylor The Holocene and Pleistocene Marine Faunas Reconsidered ................................. 25 Pierre Lozouet, Alan Beu, Philippe Maestrati, Rufino Pineda & Jean-Louis Reyss Geography of Santo and of the Sanma Province ............................................................... 34 Patricia Siméoni Contents Drainage, Hydrology and Fluvial Geomorphology ............................................................ -
OF the TOWNSVILLE REGION LAKE ROSS the Beautiful Lake Ross Stores Over 200,000 Megalitres of Water and Supplies up to 80% of Townsville’S Drinking Water
BIRDS OF THE TOWNSVILLE REGION LAKE ROSS The beautiful Lake Ross stores over 200,000 megalitres of water and supplies up to 80% of Townsville’s drinking water. The Ross River Dam wall stretches 8.3km across the Ross River floodplain, providing additional flood mitigation benefit to downstream communities. The Dam’s extensive shallow margins and fringing woodlands provide habitat for over 200 species of birds. At times, the number of Australian Pelicans, Black Swans, Eurasian Coots and Hardhead ducks can run into the thousands – a magic sight to behold. The Dam is also the breeding area for the White-bellied Sea-Eagle and the Osprey. The park around the Dam and the base of the spillway are ideal habitat for bush birds. The borrow pits across the road from the dam also support a wide variety of water birds for some months after each wet season. Lake Ross and the borrow pits are located at the end of Riverway Drive, about 14km past Thuringowa Central. Birds likely to be seen include: Australasian Darter, Little Pied Cormorant, Australian Pelican, White-faced Heron, Little Egret, Eastern Great Egret, Intermediate Egret, Australian White Ibis, Royal Spoonbill, Black Kite, White-bellied Sea-Eagle, Australian Bustard, Rainbow Lorikeet, Pale-headed Rosella, Blue-winged Kookaburra, Rainbow Bee-eater, Helmeted Friarbird, Yellow Honeyeater, Brown Honeyeater, Spangled Drongo, White-bellied Cuckoo-shrike, Pied Butcherbird, Great Bowerbird, Nutmeg Mannikin, Olive-backed Sunbird. White-faced Heron ROSS RIVER The Ross River winds its way through Townsville from Ross Dam to the mouth of the river near the Townsville Port. -
Eton Range Realignment Project ATTACHMENT 2 to EPBC Ref: 2015/7552 Preliminary Documentation Residual Impact Assessment and Offset Proposal - 37
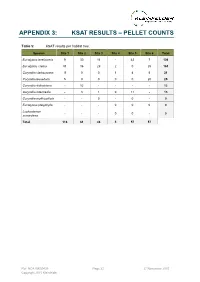
APPENDIX 3: KSAT RESULTS – PELLET COUNTS Table 5: KSAT results per habitat tree. Species Site 1 Site 2 Site 3 Site 4 Site 5 Site 6 Total Eucalyptus tereticornis 9 30 16 - 42 7 104 Eucalyptus crebra 91 16 29 2 0 25 163 Corymbia clarksoniana 11 0 0 1 4 5 21 Corymbia tessellaris 5 0 0 0 0 20 25 Corymbia dallachiana - 12 - - - - 12 Corymbia intermedia - 3 1 0 11 - 15 Corymbia erythrophloia - - 0 - 0 - 0 Eucalyptus platyphylla - - - 0 0 0 0 Lophostemon - - - 0 0 - 0 suaveolens Total 116 61 46 3 57 57 Ref: NCA15R30439 Page 22 27 November 2015 Copyright 2015 Kleinfelder APPENDIX 4: SITE PHOTOS The following images were taken from the centre of each BioCondition quadrat and represent a north east south west aspect, top left to bottom right. Ref: NCA15R30439 Page 23 27 November 2015 Copyright 2015 Kleinfelder Plate 3: BioCondition quadrat 1 (RE11.3.4/11.12.3) Ref: NCA15R30439 Page 24 27 November 2015 Copyright 2015 Kleinfelder Plate 4: BioCondition quadrat 2 (RE11.3.4/11.12.3) Ref: NCA15R30439 Page 25 27 November 2015 Copyright 2015 Kleinfelder Plate 5: BioCondition quadrat 3 (RE11.12.3) Ref: NCA15R30439 Page 26 27 November 2015 Copyright 2015 Kleinfelder Plate 6: BioCondition quadrat 4 (RE11.3.9) Ref: NCA15R30439 Page 27 27 November 2015 Copyright 2015 Kleinfelder Plate 7: BioCondition quadrat 5 (RE11.3.25) Ref: NCA15R30439 Page 28 27 November 2015 Copyright 2015 Kleinfelder Plate 8: BioCondition quadrat 6 (RE11.12.3/11.3.4/11.3.9) Ref: NCA15R30439 Page 29 27 November 2015 Copyright 2015 Kleinfelder Appendix E: Desktop Assessment for Potential