Aoteasalda and Kiwisaldula, Two New Genera of Saldidae (Hemiptera: Heteroptera), with a Key to New Zealand Genera and a New Synonymy in Zemacrosaldula
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
Schedule D Part3
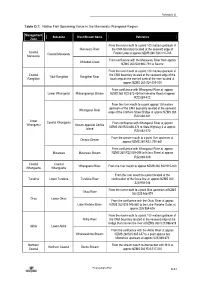
Schedule D Table D.7: Native Fish Spawning Value in the Manawatu-Wanganui Region Management Sub-zone River/Stream Name Reference Zone From the river mouth to a point 100 metres upstream of Manawatu River the CMA boundary located at the seaward edge of Coastal Coastal Manawatu Foxton Loop at approx NZMS 260 S24:010-765 Manawatu From confluence with the Manawatu River from approx Whitebait Creek NZMS 260 S24:982-791 to Source From the river mouth to a point 100 metres upstream of Coastal the CMA boundary located at the seaward edge of the Tidal Rangitikei Rangitikei River Rangitikei boat ramp on the true left bank of the river located at approx NZMS 260 S24:009-000 From confluence with Whanganui River at approx Lower Whanganui Mateongaonga Stream NZMS 260 R22:873-434 to Kaimatira Road at approx R22:889-422 From the river mouth to a point approx 100 metres upstream of the CMA boundary located at the seaward Whanganui River edge of the Cobham Street Bridge at approx NZMS 260 R22:848-381 Lower Coastal Whanganui From confluence with Whanganui River at approx Whanganui Stream opposite Corliss NZMS 260 R22:836-374 to State Highway 3 at approx Island R22:862-370 From the stream mouth to a point 1km upstream at Omapu Stream approx NZMS 260 R22: 750-441 From confluence with Whanganui River at approx Matarawa Matarawa Stream NZMS 260 R22:858-398 to Ikitara Street at approx R22:869-409 Coastal Coastal Whangaehu River From the river mouth to approx NZMS 260 S22:915-300 Whangaehu Whangaehu From the river mouth to a point located at the Turakina Lower -

Macrosaldula: Authorship and Date of Publication (Heteroptera: Saldidae)
© Zoological Institute, St.Petersburg, 2003 Macrosaldula: authorship and date of publication (Heteroptera: Saldidae) I.M. Kerzhner Kerzhner, I.M. 2003. Macrosaldula: authorship and date of publication (Heteroptera: Saldidae). Zoosystematica Rossica, 11(2), 2002: 310. The correct authorship and date for Macrosaldula is Southwood & Leston, 1959. I.M Kerzhner, Zoological Institute, Russian Academy of Sciences, Universitetskaya nab. 1, St.Petersburg 199034, Russia. E-mail: [email protected] In a key to the British Saldidae, Southwood & Leston Faraci, F. & Rizzotti Vlach, M. 1992. I Leptopodomor- (1959, p. 324) separated “Saldula (Macrosaldula) sco- pha italiani. Catalogo topografico con considerazioni tica” from “Saldula (Saldula)” by the body length and ecologiche, fenologiche e zoografiche. Mem. Soc. en- the length ratio of the second antennal segment to the tomol. Ital., 70(1991): 33-102. first one; they stated (p. 330): “S. scotica is placed in Faraci, R. & Rizzotti Vlach, M. 1995. Heteroptera. In: subgenus Macrosaldula, other British Saldula being mem- Minelli, A. et al. (Eds). Checklist delle specie della bers of the typical subgenus”. As the authors doubted fauna italiana, 41: 1-56. Bologna: Calderini. whether the name Macrosaldula was available from this Hoberlandt, L. 1977. Enumeratio insectorum Bohemo- publication, they (Leston & Southwood, 1964) published slovakiae [1]: Heteroptera. Acta faun. entomol. Mus. a special paper describing the subgenus as new with a nat. Pragae, Suppl. 4: 61-82. formal diagnosis, designation of S. scotica (Curtis) as the Leston, D. & Southwood, T.R.E. 1964. A new subgenus type species, and brief discussion mentioning the 1959 of Saldula Van Duzee, 1914 (Hem., Saldidae). Ento- publication and the acceptance of the name Macrosaldula mol. -

The Semiaquatic Hemiptera of Minnesota (Hemiptera: Heteroptera) Donald V
The Semiaquatic Hemiptera of Minnesota (Hemiptera: Heteroptera) Donald V. Bennett Edwin F. Cook Technical Bulletin 332-1981 Agricultural Experiment Station University of Minnesota St. Paul, Minnesota 55108 CONTENTS PAGE Introduction ...................................3 Key to Adults of Nearctic Families of Semiaquatic Hemiptera ................... 6 Family Saldidae-Shore Bugs ............... 7 Family Mesoveliidae-Water Treaders .......18 Family Hebridae-Velvet Water Bugs .......20 Family Hydrometridae-Marsh Treaders, Water Measurers ...22 Family Veliidae-Small Water striders, Rime bugs ................24 Family Gerridae-Water striders, Pond skaters, Wherry men .....29 Family Ochteridae-Velvety Shore Bugs ....35 Family Gelastocoridae-Toad Bugs ..........36 Literature Cited ..............................37 Figures ......................................44 Maps .........................................55 Index to Scientific Names ....................59 Acknowledgement Sincere appreciation is expressed to the following individuals: R. T. Schuh, for being extremely helpful in reviewing the section on Saldidae, lending specimens, and allowing use of his illustrations of Saldidae; C. L. Smith for reading the section on Veliidae, checking identifications, and advising on problems in the taxon omy ofthe Veliidae; D. M. Calabrese, for reviewing the section on the Gerridae and making helpful sugges tions; J. T. Polhemus, for advising on taxonomic prob lems and checking identifications for several families; C. W. Schaefer, for providing advice and editorial com ment; Y. A. Popov, for sending a copy ofhis book on the Nepomorpha; and M. C. Parsons, for supplying its English translation. The University of Minnesota, including the Agricultural Experi ment Station, is committed to the policy that all persons shall have equal access to its programs, facilities, and employment without regard to race, creed, color, sex, national origin, or handicap. The information given in this publication is for educational purposes only. -

Terrestrial Arthropod Surveys on Pagan Island, Northern Marianas
Terrestrial Arthropod Surveys on Pagan Island, Northern Marianas Neal L. Evenhuis, Lucius G. Eldredge, Keith T. Arakaki, Darcy Oishi, Janis N. Garcia & William P. Haines Pacific Biological Survey, Bishop Museum, Honolulu, Hawaii 96817 Final Report November 2010 Prepared for: U.S. Fish and Wildlife Service, Pacific Islands Fish & Wildlife Office Honolulu, Hawaii Evenhuis et al. — Pagan Island Arthropod Survey 2 BISHOP MUSEUM The State Museum of Natural and Cultural History 1525 Bernice Street Honolulu, Hawai’i 96817–2704, USA Copyright© 2010 Bishop Museum All Rights Reserved Printed in the United States of America Contribution No. 2010-015 to the Pacific Biological Survey Evenhuis et al. — Pagan Island Arthropod Survey 3 TABLE OF CONTENTS Executive Summary ......................................................................................................... 5 Background ..................................................................................................................... 7 General History .............................................................................................................. 10 Previous Expeditions to Pagan Surveying Terrestrial Arthropods ................................ 12 Current Survey and List of Collecting Sites .................................................................. 18 Sampling Methods ......................................................................................................... 25 Survey Results .............................................................................................................. -

Insecta: Hemiptera: Heteroptera)
© Zool.-Bot. Ges. Österreich, Austria; download unter www.zobodat.at Acta ZooBot Austria 155, 2018, 251–256 Snapshot of the terrestrial true bug fauna of the Pocem floodplains (Insecta: Hemiptera: Heteroptera) Wolfgang Rabitsch 61 terrestrial true bug (Heteroptera) species are reported from a short field trip in April 2017 along the river Vjosa in the Pocem floodplains, Albania. Five species are reported for the first time for Albania, indicating insufficient baseline information on the distribution of true bugs in the region. Future sampling designs should consider the interstitial habitats, river gravel and sand banks, and adjacent dry grassland areas. Certain difficulties aside, true bugs are a significant group of insect species with de- scriptive and indicative value. RABITSCH W., 2018: Momentaufnahme der terrestrischen Wanzenfauna im Pocem Überschwemmungsgebiet (Insecta: Hemiptera: Heteroptera). Während einer kurzen Exkursion im April 2017 entlang des Flusses Vjosa im Pocem Überschwemmungsgebiet, Albanien, wurden 61 terrestrische Wanzenarten (Heterop- tera) festgestellt. Fünf Arten werden der erste Mal für Albanien gemeldet, ein Hinweis auf die unzureichende Datenlage der Verbreitung von Wanzen im Gebiet. Zukünftige Erhebungen sollten insbesondere die interstitiellen Habitate am Flussufer, Sandbänke und die angrenzenden Trockenrasenstandorte untersuchen. Trotz gewisser Schwierig- keiten sind Wanzen als Deskriptoren und Indikatoren der Lebensräume eine aussage- kräftige Insektengruppe von hohem Wert. Keywords: Albania, floodplain, Heteroptera, new records, riparian, river. Introduction True bugs (Insecta: Hemiptera: Heteroptera) are well-known descriptors and indicators of terrestrial and aquatic habitats and their ecological quality (Duelli & Obrist 1998, Achtziger et al. 2007, Rabitsch 2008, Skern et al. 2010). Due to their diversity of feeding habits, life-histories, biology, and preferred habitats, true bugs are a meaningful addition to any Environmental Impact Assessment (e.g. -

Environmental Scan
Environmental Scan March 2020 www.mdc.govt.nz Environmental Scan 2020 1 Contents INTRODUCTION 5 SOCIAL AND CULTURAL PROFILE 11 ECONOMIC PROFILE 21 ENVIRONMENTAL PROFILE 31 MAJOR REGIONAL DEVELOPMENTS/PROJECTS 37 GOVERNMENT PROPOSALS, LEGISLATION, 39 INQUIRIES AND NATIONAL TRENDS BIBLIOGRAPHY 60 2 Environmental Scan 2020 Environmental Scan 2020 3 Introduction An Environmental Scan looks at what changes are likely to affect the future internal and external operating environment for Manawatū District Council (Council). It looks at where the community is heading and what we, as Council, should be doing about it. It should lead to a discussion with elected members about what tools Council has available to influence the direction the community is taking. The purpose of local government, as set out in the Local Government Act 2002 includes reference to the role of local authorities in promoting the social, economic, environmental and cultural wellbeing of their communities. The indicators included in this report have been grouped into each of the wellbeings under the headings of “Social and Cultural Profile,” “Economic Profile” and “Environmental Profile.” However, it is recognised that the many of these indicators have impacts across multiple wellbeings. Council has used the most up-to-date data available to prepare this Environmental Scan. In some cases this data is historic trend data, sometimes it is current at the time the Environmental Scan was finalised, and in some cases Council has used data and trends to prepare future forecasts. Council does not intend to update the Environmental Scan over time, but the forecasting assumptions contained within Council’s Ten Year Plan will be continually updated up until adoption. -

Foxton/Foxton Beach/Himatangi Beach PH 06 363 6007
www.manawatustandard.co.nz Manawatu Standard Saturday, April 30, 2011 61 licensed under the REAA 2008 Foxton/Foxton Beach/Himatangi Beach PH 06 363 6007 www.uniquerealty.co.nz FXF16 FXJ29 FXC39 FXU25 Max Maria van der Schouw 021 711 995 027 443 0294 1 FRANCES STREET- FOXTON 3 55 JOHNSTON STREET - FOXTON 4 42 COLEY STREET - FOXTON 4 93 UNION STREET - FOXTON 7 Affordable first home or rental investment, Instructions are clear we want SOLD! Attention big families Family home with plenty to offer within walking distance to amenities 1 1 1+ 2 Heliena Saul Nigel van der Schouw Carol Marshall Viewing By Appointment Viewing By Appointment Available For Inspection Viewing By Appointment 021 118 9132 027 262 2841 027 596 2081 Heliena 021 118 9132 $99,000 0 Nigel 027 262 2841 $136,000 2 Maria 027 443 0294 $245,000 2 Maria 027 443 0294 $265,000 2 FXN48 FXL24 FXP28 FXS64 WE HAVE A WIDE RANGE OF COASTAL SECTIONS AVAILABLE FOR SALE CALL 06 363 6007 TODAY FOR MORE INFORMATION 33 & 33A NORBITON ROAD - FOXTON 3+ 11 LINKLATER AVE - FOXTON BEACH 2 11 PRATT AVE - FOXTON BEACH 2 2 SEABURY AVE - FOXTON BEACH 3 Family home + nearly 2 acres on 2 titles in Within your reach and options of freeholding Foxton Beach bach priced at $147,000 The right time to buy is now! Ideal 1st home or town! 2 the section when it suits you 1 1 perfect investment opportunity, consider all options 1 Available For Inspection Viewing By Appointment Open Sunday 3.00-3.30pm Open Sunday 2.30-3.00pm Heliena 021 118 9132 $340,000 2 Nigel 027 262 2841 $60,000 1 Carol 027 596 2081 $147,000 -

A Review of the Hemiptera of Great Britain: the Aquatic and Semi-Aquatic Bugs
Natural England Commissioned Report NECR188 A review of the Hemiptera of Great Britain: The Aquatic and Semi-aquatic Bugs Dipsocoromorpha, Gerromorpha, Leptopodomorpha & Nepomorpha Species Status No.24 First published 20 November 2015 www.gov.uk/natural -england Foreword Natural England commission a range of reports from external contractors to provide evidence and advice to assist us in delivering our duties. The views in this report are those of the authors and do not necessarily represent those of Natural England. Background Making good decisions to conserve species should primarily be based upon an objective process of determining the degree of threat to the survival of a species. The recognised international approach to undertaking this is by assigning the species to one of the IUCN threat categories. This report was commissioned to update the national status of aquatic and semi-aquatic bugs using IUCN methodology for assessing threat. It covers all species of aquatic and semi-aquatic bugs (Heteroptera) in Great Britain, identifying those that are rare and/or under threat as well as non-threatened and non-native species. Reviews for other invertebrate groups will follow. Natural England Project Manager - Jon Webb, [email protected] Contractor - A.A. Cook (author) Keywords - invertebrates, red list, IUCN, status reviews, Heteroptera, aquatic bugs, shore bugs, IUCN threat categories, GB rarity status Further information This report can be downloaded from the Natural England website: www.gov.uk/government/organisations/natural-england. For information on Natural England publications contact the Natural England Enquiry Service on 0845 600 3078 or e-mail [email protected]. -

Microsoft Outlook
Joey Steil From: Leslie Jordan <[email protected]> Sent: Tuesday, September 25, 2018 1:13 PM To: Angela Ruberto Subject: Potential Environmental Beneficial Users of Surface Water in Your GSA Attachments: Paso Basin - County of San Luis Obispo Groundwater Sustainabilit_detail.xls; Field_Descriptions.xlsx; Freshwater_Species_Data_Sources.xls; FW_Paper_PLOSONE.pdf; FW_Paper_PLOSONE_S1.pdf; FW_Paper_PLOSONE_S2.pdf; FW_Paper_PLOSONE_S3.pdf; FW_Paper_PLOSONE_S4.pdf CALIFORNIA WATER | GROUNDWATER To: GSAs We write to provide a starting point for addressing environmental beneficial users of surface water, as required under the Sustainable Groundwater Management Act (SGMA). SGMA seeks to achieve sustainability, which is defined as the absence of several undesirable results, including “depletions of interconnected surface water that have significant and unreasonable adverse impacts on beneficial users of surface water” (Water Code §10721). The Nature Conservancy (TNC) is a science-based, nonprofit organization with a mission to conserve the lands and waters on which all life depends. Like humans, plants and animals often rely on groundwater for survival, which is why TNC helped develop, and is now helping to implement, SGMA. Earlier this year, we launched the Groundwater Resource Hub, which is an online resource intended to help make it easier and cheaper to address environmental requirements under SGMA. As a first step in addressing when depletions might have an adverse impact, The Nature Conservancy recommends identifying the beneficial users of surface water, which include environmental users. This is a critical step, as it is impossible to define “significant and unreasonable adverse impacts” without knowing what is being impacted. To make this easy, we are providing this letter and the accompanying documents as the best available science on the freshwater species within the boundary of your groundwater sustainability agency (GSA). -

A NEW GENUS of INTERTIDAL SALDIDAE from the EASTERN TROPICAL PACIFIC with NOTES on ITS BIOLOGY (Hemiptera)1
Pacific Insects ll (3-4) : 571-578 10 December 1969 A NEW GENUS OF INTERTIDAL SALDIDAE FROM THE EASTERN TROPICAL PACIFIC WITH NOTES ON ITS BIOLOGY (Hemiptera)1 By John T. Polhemus2 and William G. Evans3 Abstract: Paralosalda innova n. gen., n. sp. is described from the intertidal zone of the Pacific coast of Central America. This genus is placed in the subfamily Chiloxanthinae, and is the first known member of this group to possess 4 cells in the hemelytral mem brane. A key to the chiloxanthine genera is included, as is a summary of the intertidal saldid genera of the world, with a discussion of their relationship to Paralosalda. P. innova inhabits the mid- to upper littoral zone of protected rocky shores extending from northern Colombia to northern Costa Rica and, like other intertidal saldids, the adults spend periods of submergence by high tides in rock crevices and emerge at low tide. The nymphs, however, remain in the crevices most of the time. Until the discovery of the species described herein, only one saldid was known to ex clusively inhabit the intertidal zone in the New World : Pentacora mexicanum (Van Duzee). This insect, which evidently does not belong in Pentacora, is locally common on intertidal rocks in the northern part of the Gulf of California though the species was described from one specimen found under kelp on a beach (Van Duzee 1923). Other species of New World Saldidae are known to inhabit salt marshes and mud flats where they apparently survive submersion (for instance, Saldula notalis Drake, and Saldula palustris Douglas), but there is no evidence that they complete their life cycle in the intertidal zone or that they inhabit this zone exclusively; hence, for the present they are not considered to be intertidal in the true sense. -

Autumn 2011 Newsletter of the UK Heteroptera Recording Schemes 2Nd Series
Issue 17/18 v.1.1 Het News Autumn 2011 Newsletter of the UK Heteroptera Recording Schemes 2nd Series Circulation: An informal email newsletter circulated periodically to those interested in Heteroptera. Copyright: Text & drawings © 2011 Authors Photographs © 2011 Photographers Citation: Het News, 2nd Series, no.17/18, Spring/Autumn 2011 Editors: Our apologies for the belated publication of this year's issues, we hope that the record 30 pages in this combined issue are some compensation! Sheila Brooke: 18 Park Hill Toddington Dunstable Beds LU5 6AW — [email protected] Bernard Nau: 15 Park Hill Toddington Dunstable Beds LU5 6AW — [email protected] CONTENTS NOTICES: SOME LITERATURE ABSTRACTS ........................................... 16 Lookout for the Pondweed leafhopper ............................................................. 6 SPECIES NOTES. ................................................................18-20 Watch out for Oxycarenus lavaterae IN BRITAIN ...........................................15 Ranatra linearis, Corixa affinis, Notonecta glauca, Macrolophus spp., Contributions for next issue .................................................................................15 Conostethus venustus, Aphanus rolandri, Reduvius personatus, First incursion into Britain of Aloea australis ..................................................17 Elasmucha ferrugata Events for heteropterists .......................................................................................20 AROUND THE BRITISH ISLES............................................21-22 -
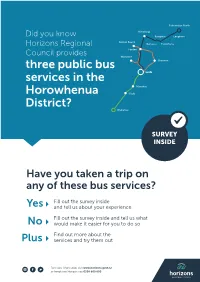
Three Public Bus Services in the Horowhenua District?
Palmerston North Himatangi Did you know Rangiotu Longburn Foxton Beach Horizons Regional Bainesse Tiakiahuna Council provides Foxton Waitarere three public bus Shannon services in the Levin Manakau Horowhenua Otaki District? Waikanae SURVEY INSIDE Have you taken a trip on any of these bus services? Fill out the survey inside Yes and tell us about your experience Fill out the survey inside and tell us what No would make it easier for you to do so Find out more about the Plus services and try them out For more information visit www.horizons.govt.nz or freephone Horizons on 0508 800 800 CURRENT TIMETABLES LEVIN TO WAIKANAE BUS SERVICE* * A minor timetable change has taken place as of 15 July 2018 Greater Wellington has made a slight adjustment to their train timetable and as result we have adjusted the bus departure times. (i.e Depart Levin changed from 9.30am to 9.20am) This operational change ensures passengers still have the opportunity to connect to the train travelling south. DEPARTS LEVIN Tuesday/Thursday Morning Time as of 15 July 2018 Location Depart Levin 9.20 am Te Takeretanga o Kura-hau-pō, Bath Street, Levin Depart Manakau 9.38 am Honi Taipua Street (opposite Manakau Bowling & Sports Club) Depart Otaki 9.51am Intercity Bus Shelter, Main Highway Otaki Arrive Waikanae 10.05am Train Station, Waikanae DEPARTS WAIKANAE Depart Waikanae 2.45 pm Train Station, Waikanae Depart Otaki 3.11 pm Intercity Bus Shelter, Main Highway Otaki Depart Manakau 3.24 pm Honi Taipua Street (opposite Manakau Bowling & Sports Club) Arrive Levin 3.38 pm