Cerambycid Pests in Forests and Urban Trees
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Download Download
March 29 2019 INSECTA 19 urn:lsid:zoobank. A Journal of World Insect Systematics org:pub:245CB278-D68A- UNDI M 4248-B9BC-8DB801F5A1EC 0695 Descriptions of one new genus and seven new species of Rhinotragini (Coleoptera: Cerambycidae: Cerambycinae) James E. Wappes American Coleoptera Museum 8734 Paisano Pass San Antonio, TX 78255-3523, USA Antonio Santos-Silva Museu de Zoologia Universidade de São Paulo São Paulo, SP, Brazil Date of issue: March 29, 2019 CENTER FOR SYSTEMATIC ENTOMOLOGY, INC., Gainesville, FL James E. Wappes and Antonio Santos-Silva Descriptions of one new genus and seven new species of Rhinotragini (Coleoptera: Cerambycidae: Cerambycinae) Insecta Mundi 0695: 1–19 ZooBank Registered: urn:lsid:zoobank.org:pub:245CB278-D68A-4248-B9BC-8DB801F5A1EC Published in 2019 by Center for Systematic Entomology, Inc. P.O. Box 141874 Gainesville, FL 32614-1874 USA http://centerforsystematicentomology.org/ Insecta Mundi is a journal primarily devoted to insect systematics, but articles can be published on any non- marine arthropod. Topics considered for publication include systematics, taxonomy, nomenclature, checklists, faunal works, and natural history. Insecta Mundi will not consider works in the applied sciences (i.e. medical entomology, pest control research, etc.), and no longer publishes book reviews or editorials. Insecta Mundi publishes original research or discoveries in an inexpensive and timely manner, distributing them free via open access on the internet on the date of publication. Insecta Mundi is referenced or abstracted by several sources, including the Zoological Record and CAB Abstracts. Insecta Mundi is published irregularly throughout the year, with completed manuscripts assigned an individual number. Manuscripts must be peer reviewed prior to submission, after which they are reviewed by the editorial board to ensure quality. -

(Coleoptera) of Peru Miguel A
University of Nebraska - Lincoln DigitalCommons@University of Nebraska - Lincoln Center for Systematic Entomology, Gainesville, Insecta Mundi Florida 2-29-2012 Preliminary checklist of the Cerambycidae, Disteniidae, and Vesperidae (Coleoptera) of Peru Miguel A. Monné Universidade Federal do Rio de Janeiro, [email protected] Eugenio H. Nearns University of New Mexico, [email protected] Sarah C. Carbonel Carril Universidad Nacional Mayor de San Marcos, Peru, [email protected] Ian P. Swift California State Collection of Arthropods, [email protected] Marcela L. Monné Universidade Federal do Rio de Janeiro, [email protected] Follow this and additional works at: http://digitalcommons.unl.edu/insectamundi Part of the Entomology Commons Monné, Miguel A.; Nearns, Eugenio H.; Carbonel Carril, Sarah C.; Swift, Ian P.; and Monné, Marcela L., "Preliminary checklist of the Cerambycidae, Disteniidae, and Vesperidae (Coleoptera) of Peru" (2012). Insecta Mundi. Paper 717. http://digitalcommons.unl.edu/insectamundi/717 This Article is brought to you for free and open access by the Center for Systematic Entomology, Gainesville, Florida at DigitalCommons@University of Nebraska - Lincoln. It has been accepted for inclusion in Insecta Mundi by an authorized administrator of DigitalCommons@University of Nebraska - Lincoln. INSECTA MUNDI A Journal of World Insect Systematics 0213 Preliminary checklist of the Cerambycidae, Disteniidae, and Vesperidae (Coleoptera) of Peru Miguel A. Monné Museu Nacional Universidade Federal do Rio de Janeiro Quinta da Boa Vista São Cristóvão, 20940-040 Rio de Janeiro, RJ, Brazil Eugenio H. Nearns Department of Biology Museum of Southwestern Biology University of New Mexico Albuquerque, NM 87131-0001, USA Sarah C. Carbonel Carril Departamento de Entomología Museo de Historia Natural Universidad Nacional Mayor de San Marcos Avenida Arenales 1256, Lima, Peru Ian P. -

Coleoptera, Cerambycidae, Lamiinae) with Description of a Species with Non‑Retractile Parameres
ARTICLE Two new genera of Desmiphorini (Coleoptera, Cerambycidae, Lamiinae) with description of a species with non‑retractile parameres Francisco Eriberto de Lima Nascimento¹² & Antonio Santos-Silva¹³ ¹ Universidade de São Paulo (USP), Museu de Zoologia (MZUSP). São Paulo, SP, Brasil. ² ORCID: http://orcid.org/0000-0002-5047-8921. E-mail: [email protected] ³ ORCID: http://orcid.org/0000-0001-7128-1418. E-mail: [email protected] Abstract. In this study, two new genera of Desmiphorini (Lamiinae) are proposed: Cleidaria gen. nov., to include Cleidaria cleidae sp. nov. from the state of Chiapas in Mexico, and Obscenoides gen. nov. for Desmiphora (D.) compta Martins & Galileo, 2005. The shape of tarsal claws of Cleidaria cleidae sp. nov. (abruptly narrowed from basal half) is so far, not found in any current genera of the tribe. With respect to Obscenoides compta (Martins & Galileo, 2005) comb. nov., the genitalia of males have an unusual shape with non-retractile parameres. The character combination related to this genital structure is unknown to us in other species in the family, and hypotheses about its function are suggested. Key-Words. Genital morphology; Longhorned beetles; New taxa; Taxonomy. INTRODUCTION the current definitions of some tribes, especially based on the works of Breuning do not take into Lamiinae (Cerambycidae), also known as flat- account adaptive convergences and use superfi- faced longhorns, with more than 21,000 described cial characters to subordinate taxa. species in about 3,000 genera and 87 tribes is Among these tribes, Desmiphorini Thomson, the largest subfamily of Cerambycidae occurring 1860 is not an exception, and its “boundaries” are worldwide (Tavakilian & Chevillotte, 2019). -

Ethanol and (–)-A-Pinene: Attractant Kairomones for Some Large Wood-Boring Beetles in Southeastern USA
J Chem Ecol (2006) DOI 10.1007/s10886-006-9037-8 Ethanol and (–)-a-Pinene: Attractant Kairomones for Some Large Wood-Boring Beetles in Southeastern USA Daniel R. Miller Received: 12 September 2005 /Revised: 12 December 2005 /Accepted: 2 January 2006 # Springer Science + Business Media, Inc. 2006 Abstract Ethanol and a-pinene were tested as attractants for large wood-boring pine beetles in Alabama, Florida, Georgia, North Carolina, and South Carolina in 2002–2004. Multiple-funnel traps baited with (j)-a-pinene (released at about 2 g/d at 25–28-C) were attractive to the following Cerambycidae: Acanthocinus nodosus, A. obsoletus, Arhopalus rusticus nubilus, Asemum striatum, Monochamus titillator, Prionus pocularis, Xylotrechus integer, and X. sagittatus sagittatus. Buprestis lineata (Buprestidae), Alaus myops (Elateridae), and Hylobius pales and Pachylobius picivorus (Curculionidae) were also attracted to traps baited with (j)-a-pinene. In many locations, ethanol synergized attraction of the cerambycids Acanthocinus nodosus, A. obsoletus, Arhopalus r. nubilus, Monochamus titillator, and Xylotrechus s. sagittatus (but not Asemum striatum, Prionus pocularis,orXylotrechus integer)to traps baited with (j)-a-pinene. Similarly, attraction of Alaus myops, Hylobius pales, and Pachylobius picivorus (but not Buprestis lineata) to traps baited with (j)-a- pinene was synergized by ethanol. These results provide support for the use of traps baited with ethanol and (j)-a-pinene to detect and monitor common large wood- boring beetles from the southeastern region of the USA at ports-of-entry in other countries, as well as forested areas in the USA. Keywords Cerambycidae . Xylotrechus . Monochamus . Acanthocinus Curculionidae . Hylobius . Pachylobius . Elateridae . Alaus . Ethanol a-Pinene . -
Insect Pest List by Host Tree and Reported Country
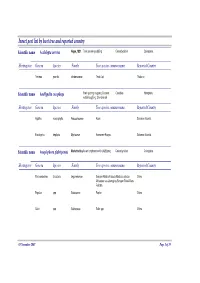
Insect pest list by host tree and reported country Scientific name Acalolepta cervina Hope, 1831 Teak canker grub|Eng Cerambycidae Coleoptera Hosting tree Genera Species Family Tree species common name Reported Country Tectona grandis Verbenaceae Teak-Jati Thailand Scientific name Amblypelta cocophaga Fruit spotting bug|eng Coconut Coreidae Hemiptera nutfall bug|Eng, Chinche del Hosting tree Genera Species Family Tree species common name Reported Country Agathis macrophylla Araucariaceae Kauri Solomon Islands Eucalyptus deglupta Myrtaceae Kamarere-Bagras Solomon Islands Scientific name Anoplophora glabripennis Motschulsky Asian longhorn beetle (ALB)|eng Cerambycidae Coleoptera Hosting tree Genera Species Family Tree species common name Reported Country Paraserianthes falcataria Leguminosae Sengon-Albizia-Falcata-Molucca albizia- China Moluccac sau-Jeungjing-Sengon-Batai-Mara- Falcata Populus spp. Salicaceae Poplar China Salix spp. Salicaceae Salix spp. China 05 November 2007 Page 1 of 35 Scientific name Aonidiella orientalis Newstead, Oriental scale|eng Diaspididae Homoptera 1894 Hosting tree Genera Species Family Tree species common name Reported Country Lovoa swynnertonii Meliaceae East African walnut Cameroon Azadirachta indica Meliaceae Melia indica-Neem Nigeria Scientific name Apethymus abdominalis Lepeletier, Tenthredinidae Hymenoptera 1823 Hosting tree Genera Species Family Tree species common name Reported Country Other Coniferous Other Coniferous Romania Scientific name Apriona germari Hope 1831 Long-horned beetle|eng Cerambycidae -

0865 a New Genus Cicatrisphaerion, New Species, New Records, Page Count: 20 and Redescriptions of Neotropical Cerambycidae (Coleoptera) Steven W
A journal of world insect systematics INSECTA MUNDI 0865 A new genus Cicatrisphaerion, new species, new records, Page Count: 20 and redescriptions of Neotropical Cerambycidae (Coleoptera) Steven W. Lingafelter Lingafelter et al. 8920 S. Bryerly Ct. Hereford, Arizona 85615, USA. Roy F. Morris II 2635 Ewell Road Lakeland, Florida 33811, USA [email protected] Frederick W. Skillman Jr. 7033 S. 19th St. Phoenix, Arizona 85042, USA Antonio Santos-Silva Museu de Zoologia, Universidade de São Paulo São Paulo, SP, Brazil Date of issue: April 30, 2021 Center for Systematic Entomology, Inc., Gainesville, FL Lingafelter SW, Morris RF II, Skillman FW Jr., Santos-Silva A. 2021. A new genus Cicatrisphaerion, new species, new records, and redescriptions of Neotropical Cerambycidae (Coleoptera). Insecta Mundi 0865: 1–20. Published on April 30, 2021 by Center for Systematic Entomology, Inc. P.O. Box 141874 Gainesville, FL 32614-1874 USA http://centerforsystematicentomology.org/ Insecta Mundi is a journal primarily devoted to insect systematics, but articles can be published on any non- marine arthropod. Topics considered for publication include systematics, taxonomy, nomenclature, checklists, faunal works, and natural history. Insecta Mundi will not consider works in the applied sciences (i.e. medi- cal entomology, pest control research, etc.), and no longer publishes book reviews or editorials. Insecta Mundi publishes original research or discoveries in an inexpensive and timely manner, distributing them free via open access on the internet on the date of publication. Insecta Mundi is referenced or abstracted by several sources, including the Zoological Record and CAB Abstracts. Insecta Mundi is published irregularly throughout the year, with completed manuscripts assigned an individual number. -

Your Name Here
RELATIONSHIPS BETWEEN DEAD WOOD AND ARTHROPODS IN THE SOUTHEASTERN UNITED STATES by MICHAEL DARRAGH ULYSHEN (Under the Direction of James L. Hanula) ABSTRACT The importance of dead wood to maintaining forest diversity is now widely recognized. However, the habitat associations and sensitivities of many species associated with dead wood remain unknown, making it difficult to develop conservation plans for managed forests. The purpose of this research, conducted on the upper coastal plain of South Carolina, was to better understand the relationships between dead wood and arthropods in the southeastern United States. In a comparison of forest types, more beetle species emerged from logs collected in upland pine-dominated stands than in bottomland hardwood forests. This difference was most pronounced for Quercus nigra L., a species of tree uncommon in upland forests. In a comparison of wood postures, more beetle species emerged from logs than from snags, but a number of species appear to be dependent on snags including several canopy specialists. In a study of saproxylic beetle succession, species richness peaked within the first year of death and declined steadily thereafter. However, a number of species appear to be dependent on highly decayed logs, underscoring the importance of protecting wood at all stages of decay. In a study comparing litter-dwelling arthropod abundance at different distances from dead wood, arthropods were more abundant near dead wood than away from it. In another study, ground- dwelling arthropods and saproxylic beetles were little affected by large-scale manipulations of dead wood in upland pine-dominated forests, possibly due to the suitability of the forests surrounding the plots. -

(Coleoptera) of the Huron Mountains in Northern Michigan
The Great Lakes Entomologist Volume 19 Number 3 - Fall 1986 Number 3 - Fall 1986 Article 3 October 1986 Ecology of the Cerambycidae (Coleoptera) of the Huron Mountains in Northern Michigan D. C. L. Gosling Follow this and additional works at: https://scholar.valpo.edu/tgle Part of the Entomology Commons Recommended Citation Gosling, D. C. L. 1986. "Ecology of the Cerambycidae (Coleoptera) of the Huron Mountains in Northern Michigan," The Great Lakes Entomologist, vol 19 (3) Available at: https://scholar.valpo.edu/tgle/vol19/iss3/3 This Peer-Review Article is brought to you for free and open access by the Department of Biology at ValpoScholar. It has been accepted for inclusion in The Great Lakes Entomologist by an authorized administrator of ValpoScholar. For more information, please contact a ValpoScholar staff member at [email protected]. Gosling: Ecology of the Cerambycidae (Coleoptera) of the Huron Mountains i 1986 THE GREAT LAKES ENTOMOLOGIST 153 ECOLOGY OF THE CERAMBYCIDAE (COLEOPTERA) OF THE HURON MOUNTAINS IN NORTHERN MICHIGAN D. C. L Gosling! ABSTRACT Eighty-nine species of Cerambycidae were collected during a five-year survey of the woodboring beetle fauna of the Huron Mountains in Marquette County, Michigan. Host plants were deteTITIined for 51 species. Observations were made of species abundance and phenology, and the blossoms visited by anthophilous cerambycids. The Huron Mountains area comprises approximately 13,000 ha of forested land in northern Marquette County in the Upper Peninsula of Michigan. More than 7000 ha are privately owned by the Huron Mountain Club, including a designated, 2200 ha, Nature Research Area. The variety of habitats combines with differences in the nature and extent of prior disturbance to produce an exceptional diversity of forest communities, making the area particularly valuable for studies of forest insects. -

Ramesh Chander Bhagat.Pdf
Int. J. Curr. Res. Biosci. Plant Biol. 2016, 3(12): 115-123 International Journal of Current Research in Biosciences and Plant Biology ISSN: 2349-8080 (Online) ● Volume 3 ● Number 12 (December-2016) Journal homepage: www.ijcrbp.com Original Research Article doi: http://dx.doi.org/10.20546/ijcrbp.2016.312.014 An Update on Checklist and Biodiversity of Coleopteran-fauna (Insecta) of Forestry and Mulberry Importance in Jammu and Kashmir State (India) Ramesh Chander Bhagat* P.O. Box No. 1250, G.P.O., Residency Road, Srinagar, Kashmir-190 001, J & K, India *Corresponding author. A b s t r a c t Article Info The present paper deals with a total of 64 species of beetles and weevils (Coleoptera), Accepted: 29 November 2016 belonging to 52 genera, under 14 families, associated with diverse species of forest and Available Online: 06 December 2016 mulberry plantations, occurring in vast areas and localities of Jammu and Kashmir State. The Coleopteran species of forestry and mulberry importance accounts for 73.43 K e y w o r d s % and 35.93 % respectively. The Coleopteran-fauna (47 spp.), spread over 12 families, is found to be infesting forest trees,viz. Ash, Benne, Birch, Conifers, Elms, Biodiversity Ivy, Maple, Oak, Parrotia, Plane tree, Poplars, Robinia, Salix, and Yew. Of these trees, Checklist Pines showed highest number of Coleopteran species i.e. 18, under 6 families, followed Coleopteran-fauna by Poplars, with 15 spp. (4 families) and Cedars, having 14 spp. (4 families) The Forest trees Mulberry plantations Mulberry plantations (Morus spp.) both endemic as well as exotic, have been observed to be infesting 23 spp. -

Samano Et Al 2012 Monochamus Pheromones CE
821 2-Undecyloxy-1-ethanol in combination with other semiochemicals attracts three Monochamus species (Coleoptera: Cerambycidae) in British Columbia, Canada Jorge E. Macias-Samano,1 David Wakarchuk, Jocelyn G. Millar, Lawrence M. Hanks Abstract—Two species in the genus Monochamus Dejean (Coleoptera: Cerambycidae: Lamiinae) have recently been shown to have the same male-produced sex pheromone, 2-undecyloxy-1-ethanol (monochamol), suggesting that other congeners may share the same pheromone. We tested that hypothesis by conducting field bioassays of monochamol, in combination with bark-beetle pheromones and the host plant volatiles ethanol and a-pinene, in southern British Columbia, Canada. We captured 603 Monochamus clamator (LeConte), 63 Monochamus obtusus Casey, 245 Monochamus scutellatus (Say) (tribe Monochamini), and 42 Acanthocinus princeps (Walker) (tribe Acanthocinini). All three Mono- chamus species were significantly attracted to the combination of monochamol and host plant volatiles, whereas bark-beetle pheromones plus plant volatiles and plant volatiles alone were minimally attractive. Adding bark-beetle pheromones to the monochamol plus plant volatiles treatment synergised attraction of M. clamator, but not the other two Monochamus species. Acanthocinus princeps was most strongly attracted to the combination of bark-beetle pheromones and plant volatiles, and did not appear to be affected by the presence or absence of monochamol in baits. We conclude that monochamol is a likely pheromone component for the three Monochamus species, and that monochamol plus host plant volatiles is an effective attractant for these and perhaps other North American Monochamus species. Re´sume´—On a de´montre´re´cemment que deux espe`ces du genre Monochamus Dejean (Coleoptera: Cerambycidae: Lamiinae) posse´daient la meˆme phe´romone sexuelle maˆle, le 2-unde´cyloxy-1-e´thanol (monochamol), ce qui laisse croire que d’autres conge´ne`res pourraient aussi avoir cette meˆme phe´romone. -

Butterflies of North America
Insects of Western North America 7. Survey of Selected Arthropod Taxa of Fort Sill, Comanche County, Oklahoma. 4. Hexapoda: Selected Coleoptera and Diptera with cumulative list of Arthropoda and additional taxa Contributions of the C.P. Gillette Museum of Arthropod Diversity Colorado State University, Fort Collins, CO 80523-1177 2 Insects of Western North America. 7. Survey of Selected Arthropod Taxa of Fort Sill, Comanche County, Oklahoma. 4. Hexapoda: Selected Coleoptera and Diptera with cumulative list of Arthropoda and additional taxa by Boris C. Kondratieff, Luke Myers, and Whitney S. Cranshaw C.P. Gillette Museum of Arthropod Diversity Department of Bioagricultural Sciences and Pest Management Colorado State University, Fort Collins, Colorado 80523 August 22, 2011 Contributions of the C.P. Gillette Museum of Arthropod Diversity. Department of Bioagricultural Sciences and Pest Management Colorado State University, Fort Collins, CO 80523-1177 3 Cover Photo Credits: Whitney S. Cranshaw. Females of the blow fly Cochliomyia macellaria (Fab.) laying eggs on an animal carcass on Fort Sill, Oklahoma. ISBN 1084-8819 This publication and others in the series may be ordered from the C.P. Gillette Museum of Arthropod Diversity, Department of Bioagricultural Sciences and Pest Management, Colorado State University, Fort Collins, Colorado, 80523-1177. Copyrighted 2011 4 Contents EXECUTIVE SUMMARY .............................................................................................................7 SUMMARY AND MANAGEMENT CONSIDERATIONS -

PRA on Apriona Species
EUROPEAN AND MEDITERRANEAN PLANT PROTECTION ORGANIZATION ORGANISATION EUROPEENNE ET MEDITERRANEENNE POUR LA PROTECTION DES PLANTES 16-22171 (13-18692) Only the yellow note is new compared to document 13-18692 Pest Risk Analysis for Apriona germari, A. japonica, A. cinerea Note: This PRA started 2011; as a result, three species of Apriona were added to the EPPO A1 List: Apriona germari, A. japonica and A. cinerea. However recent taxonomic changes have occurred with significant consequences on their geographical distributions. A. rugicollis is no longer considered as a synonym of A. germari but as a distinct species. A. japonica, which was previously considered to be a distinct species, has been synonymized with A. rugicollis. Finally, A. cinerea remains a separate species. Most of the interceptions reported in the EU as A. germari are in fact A. rugicollis. The outcomes of the PRA for these pests do not change. However A. germari has a more limited and a more tropical distribution than originally assessed, but it is considered that it could establish in Southern EPPO countries. The Panel on Phytosanitary Measures agreed with the addition of Apriona rugicollis to the A1 list. September 2013 EPPO 21 Boulevard Richard Lenoir 75011 Paris www.eppo.int [email protected] This risk assessment follows the EPPO Standard PM PM 5/3(5) Decision-support scheme for quarantine pests (available at http://archives.eppo.int/EPPOStandards/pra.htm) and uses the terminology defined in ISPM 5 Glossary of Phytosanitary Terms (available at https://www.ippc.int/index.php). This document was first elaborated by an Expert Working Group and then reviewed by the Panel on Phytosanitary Measures and if relevant other EPPO bodies.