*Scottish Birds 25
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

2019 Cruise Directory
Despite the modern fashion for large floating resorts, we b 7 nights 0 2019 CRUISE DIRECTORY Highlands and Islands of Scotland Orkney and Shetland Northern Ireland and The Isle of Man Cape Wrath Scrabster SCOTLAND Kinlochbervie Wick and IRELAND HANDA ISLAND Loch a’ FLANNAN Stornoway Chàirn Bhain ISLES LEWIS Lochinver SUMMER ISLES NORTH SHIANT ISLES ST KILDA Tarbert SEA Ullapool HARRIS Loch Ewe Loch Broom BERNERAY Trotternish Inverewe ATLANTIC NORTH Peninsula Inner Gairloch OCEAN UIST North INVERGORDON Minch Sound Lochmaddy Uig Shieldaig BENBECULA Dunvegan RAASAY INVERNESS SKYE Portree Loch Carron Loch Harport Kyle of Plockton SOUTH Lochalsh UIST Lochboisdale Loch Coruisk Little Minch Loch Hourn ERISKAY CANNA Armadale BARRA RUM Inverie Castlebay Sound of VATERSAY Sleat SCOTLAND PABBAY EIGG MINGULAY MUCK Fort William BARRA HEAD Sea of the Glenmore Loch Linnhe Hebrides Kilchoan Bay Salen CARNA Ballachulish COLL Sound Loch Sunart Tobermory Loch à Choire TIREE ULVA of Mull MULL ISLE OF ERISKA LUNGA Craignure Dunsta!nage STAFFA OBAN IONA KERRERA Firth of Lorn Craobh Haven Inveraray Ardfern Strachur Crarae Loch Goil COLONSAY Crinan Loch Loch Long Tayvallich Rhu LochStriven Fyne Holy Loch JURA GREENOCK Loch na Mile Tarbert Portavadie GLASGOW ISLAY Rothesay BUTE Largs GIGHA GREAT CUMBRAE Port Ellen Lochranza LITTLE CUMBRAE Brodick HOLY Troon ISLE ARRAN Campbeltown Firth of Clyde RATHLIN ISLAND SANDA ISLAND AILSA Ballycastle CRAIG North Channel NORTHERN Larne IRELAND Bangor ENGLAND BELFAST Strangford Lough IRISH SEA ISLE OF MAN EIRE Peel Douglas ORKNEY and Muckle Flugga UNST SHETLAND Baltasound YELL Burravoe Lunna Voe WHALSAY SHETLAND Lerwick Scalloway BRESSAY Grutness FAIR ISLE ATLANTIC OCEAN WESTRAY SANDAY STRONSAY ORKNEY Kirkwall Stromness Scapa Flow HOY Lyness SOUTH RONALDSAY NORTH SEA Pentland Firth STROMA Scrabster Caithness Wick Welcome to the 2019 Hebridean Princess Cruise Directory Unlike most cruise companies, Hebridean operates just one very small and special ship – Hebridean Princess. -

2020 Cruise Directory Directory 2020 Cruise 2020 Cruise Directory M 18 C B Y 80 −−−−−−−−−−−−−−− 17 −−−−−−−−−−−−−−−
2020 MAIN Cover Artwork.qxp_Layout 1 07/03/2019 16:16 Page 1 2020 Hebridean Princess Cruise Calendar SPRING page CONTENTS March 2nd A Taste of the Lower Clyde 4 nights 22 European River Cruises on board MS Royal Crown 6th Firth of Clyde Explorer 4 nights 24 10th Historic Houses and Castles of the Clyde 7 nights 26 The Hebridean difference 3 Private charters 17 17th Inlets and Islands of Argyll 7 nights 28 24th Highland and Island Discovery 7 nights 30 Genuinely fully-inclusive cruising 4-5 Belmond Royal Scotsman 17 31st Flavours of the Hebrides 7 nights 32 Discovering more with Scottish islands A-Z 18-21 Hebridean’s exceptional crew 6-7 April 7th Easter Explorer 7 nights 34 Cruise itineraries 22-97 Life on board 8-9 14th Springtime Surprise 7 nights 36 Cabins 98-107 21st Idyllic Outer Isles 7 nights 38 Dining and cuisine 10-11 28th Footloose through the Inner Sound 7 nights 40 Smooth start to your cruise 108-109 2020 Cruise DireCTOrY Going ashore 12-13 On board A-Z 111 May 5th Glorious Gardens of the West Coast 7 nights 42 Themed cruises 14 12th Western Isles Panorama 7 nights 44 Highlands and islands of scotland What you need to know 112 Enriching guest speakers 15 19th St Kilda and the Outer Isles 7 nights 46 Orkney, Northern ireland, isle of Man and Norway Cabin facilities 113 26th Western Isles Wildlife 7 nights 48 Knowledgeable guides 15 Deck plans 114 SuMMER Partnerships 16 June 2nd St Kilda & Scotland’s Remote Archipelagos 7 nights 50 9th Heart of the Hebrides 7 nights 52 16th Footloose to the Outer Isles 7 nights 54 HEBRIDEAN -

Where to Go: Puffin Colonies in Ireland Over 15,000 Puffin Pairs Were Recorded in Ireland at the Time of the Last Census
Where to go: puffin colonies in Ireland Over 15,000 puffin pairs were recorded in Ireland at the time of the last census. We are interested in receiving your photos from ANY colony and the grid references for known puffin locations are given in the table. The largest and most accessible colonies here are Great Skellig and Great Saltee. Start Number Site Access for Pufferazzi Further information Grid of pairs Access possible for Puffarazzi, but Great Skellig V247607 4,000 worldheritageireland.ie/skellig-michael check local access arrangements Puffin Island - Kerry V336674 3,000 Access more difficult Boat trips available but landing not possible 1,522 Access possible for Puffarazzi, but Great Saltee X950970 salteeislands.info check local access arrangements Mayo Islands l550938 1,500 Access more difficult Illanmaster F930427 1,355 Access more difficult Access possible for Puffarazzi, but Cliffs of Moher, SPA R034913 1,075 check local access arrangements Stags of Broadhaven F840480 1,000 Access more difficult Tory Island and Bloody B878455 894 Access more difficult Foreland Kid Island F785435 370 Access more difficult Little Saltee - Wexford X968994 300 Access more difficult Inishvickillane V208917 170 Access more difficult Access possible for Puffarazzi, but Horn Head C005413 150 check local access arrangements Lambay Island O316514 87 Access more difficult Pig Island F880437 85 Access more difficult Inishturk Island L594748 80 Access more difficult Clare Island L652856 25 Access more difficult Beldog Harbour to Kid F785435 21 Access more difficult Island Mayo: North West F483156 7 Access more difficult Islands Ireland’s Eye O285414 4 Access more difficult Howth Head O299389 2 Access more difficult Wicklow Head T344925 1 Access more difficult Where to go: puffin colonies in Inner Hebrides Over 2,000 puffin pairs were recorded in the Inner Hebrides at the time of the last census. -

Hiking Scotland's
Hiking Scotland’s North Highlands & Isle of Lewis July 20-30, 2021 (11 days | 15 guests) with archaeologist Mary MacLeod Rivett Archaeology-focused tours for the curious to the connoisseur. Clachtoll Broch Handa Island Arnol Dun Carloway (5.5|645) (6|890) BORVE Great Bernera & Traigh Uige 3 Caithness Dunbeath(4.5|425) (6|870) Stornoway (5|~) 3 3 BRORA Glasgow Isle of Lewis Callanish Lairg Standing Stones Ullapool (4.5|~) (4.5|885) Ardvreck LOCHINVER Castle Inverness Little Assynt # Overnight stays Itinerary stops Scottish Flights Hikes (miles|feet) Highlands Ferry Archaeological Institute of America Lecturer & Host Dr. Mary MacLeod oin archaeologist Mary MacLeod Rivett and a small group of like- Rivett was born in minded travelers on this 11-day tour of Scotland’s remote north London, England, to J Highlands and the Isle of Lewis in the Outer Hebrides. Mostly we a Scottish-Canadian family. Her father’s will explore off the well-beaten Highland tourist trail, and along the way family was from we will be treated to an abundance of archaeological and historical sites, Scotland’s Outer striking scenery – including high cliffs, sea lochs, sandy and rocky bays, Hebrides, and she mountains, and glens – and, of course, excellent hiking. spent a lot of time in the Hebrides as a child. Mary earned her Scotland’s long and varied history stretches back many thousands of B.A. from the University of Cambridge, years, and archaeological remains ranging from Neolithic cairns and and her M.A. from the University of stone circles to Iron Age brochs (ancient dry stone buildings unique to York. -
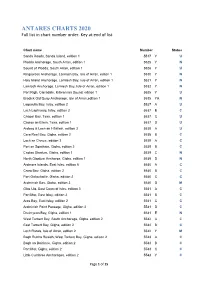
ANTARES CHARTS 2020 Full List in Chart Number Order
ANTARES CHARTS 2020 Full list in chart number order. Key at end of list Chart name Number Status Sanda Roads, Sanda Island, edition 1 5517 Y U Pladda Anchorage, South Arran, edition 1 5525 Y N Sound of Pladda, South Arran, edition 1 5526 Y U Kingscross Anchorage, Lamlash Bay, Isle of Arran, editon 1 5530 Y N Holy Island Anchorage, Lamlash Bay, Isle of Arran, edition 1 5531 Y N Lamlash Anchorage, Lamlash Bay, Isle of Arran, edition 1 5532 Y N Port Righ, Carradale, Kilbrannan Sound, edition 1 5535 Y U Brodick Old Quay Anchorage, Isle of Arran,edition 1 5535 YA N Lagavulin Bay, Islay, edition 2 5537 A U Loch Laphroaig, Islay, edition 2 5537 B C Chapel Bay, Texa, edition 1 5537 C U Caolas an Eilein, Texa, edition 1 5537 D U Ardbeg & Loch an t-Sailein, edition 3 5538 A U Cara Reef Bay, Gigha, edition 2 5538 B C Loch an Chnuic, edition 3 5539 A C Port an Sgiathain, Gigha, edition 2 5539 B C Caolas Gigalum, Gigha, edition 1 5539 C N North Gigalum Anchorge, Gigha, edition 1 5539 D N Ardmore Islands, East Islay, edition 5 5540 A C Craro Bay, Gigha, edition 2 5540 B C Port Gallochoille, Gigha, edition 2 5540 C C Ardminish Bay, Gigha, edition 3 5540 D M Glas Uig, East Coast of Islay, edition 3 5541 A C Port Mor, East Islay, edition 2 5541 B C Aros Bay, East Islay, edition 2 5541 C C Ardminish Point Passage, Gigha, edition 2 5541 D C Druimyeon Bay, Gigha, edition 1 5541 E N West Tarbert Bay, South Anchorage, Gigha, edition 2 5542 A C East Tarbert Bay, Gigha, edition 2 5542 B C Loch Ranza, Isle of Arran, edition 2 5542 Y M Bagh Rubha Ruaidh, West Tarbert -

Scottish Sea Kayaking Sea Scottish
Scottish Sea Kayaking At last, here it is… Scotland’s first guidebook for sea kayakers wishing to explore its amazing coastline and magical islands. It brings together a selection of fifty great sea voyages around the mainland of Scotland, Doug Cooper & George Reid from the Mull of Galloway in the SW to St Abb’s Head on the east coast, as well as voyages in the Western Isles, ranging from day trips to three day journeys. Illustrated with superb colour photographs and useful maps throughout, it is a practical guide to help you select and plan trips. It will provide inspiration for future voyages and a souvenir of journeys undertaken. As well as providing essential information on where to start and finish, distances, times and tidal information, the book does much to stimulate and inform our interest in the environment we are passing through. It is full of facts and anecdotes about local history, geology, scenery, seabirds and sea mammals. A fascinating read and an inspirational book. Scottish Sea Kayaking Scottish Sea Kayaking fifty great sea kayak voyages fifty great sea kayak voyages Doug Cooper & George Reid Also available from 35 36 38 39 33 37 Pesda Press 40 27 26 Sea Kayak Navigation 41 The Seamanship Pocketbook Stornoway 32 25 Welsh Sea Kayaking 24 South West Sea Kayaking 22 34 21 The Northern Isles 31 23 Oileáin - A Guide to the Irish Islands 18 43 30 17 44 Kayak Surfing 42 16 Kayak Rolling 29 19 Scottish White Water 15 Inverness 45 English White Water 14 20 Canoe and Kayak Handbook White Water Safety and Rescue 13 .. -

Rattus Norvegicus) from the Isle of Canna, Inner Hebrides, Scotland
Bell, E.; D. Boyle, K. Floyd, P. Garner-Richards, B. Swann, R. Luxmoore, A. Patterson, and R. Thomas. The ground-based eradication of Norway rats (Rattus norvegicus) from the Isle of Canna, Inner Hebrides, Scotland The ground-based eradication of Norway rats (Rattus norvegicus) from the Isle of Canna, Inner Hebrides, Scotland E. Bell1, D. Boyle1, K. Floyd1, P. Garner-Richards1, B. Swann2, R. Luxmoore3, A. Patterson4, and R. Thomas5 1Wildlife Management International Limited, 35 Selmes Road, Rapaura, RD3, Blenheim, 7273, New Zealand. <[email protected]>. 214 St Vincent Road, Tain, Ross-shire, IV19 1JR, Scotland. 3Scotland. National Trust for Scotland, 28 Charlotte Square, Edinburgh, EH2 4ET, Scotland. 4ABI Wildlife Consultancy, 4 Carnbee End, Edinburgh, EH16 6GJ, Scotland. 5The Royal Zoological Society of Scotland, 134 Corstorphine Road, Edinburgh, EH12 6TS, Scotland. Abstract: Seabird populations, particularly European shags (Phalacrocorax aristotelis) and Manx shearwater (Puffinus puffinus), on the Isle of Canna have been in decline for several years. Norway rats (Rattus norvegicus) were identified as the most likely factor. The Canna Seabird Recovery Project, developed as a result of this information, was a three year project incorporating the ground-based eradication of Norway rats (Phase I) followed by long-term monitoring, quarantine and contingency for rodents (Phase II) and continued long-term monitoring of the seabird populations (Phase III). The National Trust for Scotland (NTS) contracted Wildlife Management International Limited to direct the first and second phases of the project (August 2005-June 2008), with the assistance of NTS staff and volunteers. Techniques, problems, non-target species, (particularly wood mice (Apodemus sylvaticus) and white-tailed eagles (Haliaeetus albicilla)) issues, solutions and results of the operation on the permanently inhabited island are covered. -

Hebridean Princess Is Like No Ordinary Cruise
2021 CRUISE DIRECTORY Highlands and Islands of Scotland Orkney, Northern Ireland, Isle of Man and Norway Cruising on board Hebridean Princess is like no ordinary cruise On a Hebridean Princess cruise you can relax and enjoy the journey of a lifetime, safe in the knowledge that your holiday is genuinely fully-inclusive. Cruises shown in this booklet include: • Shore visits including all entrance fees and transfers; • All meals on board and ashore, snacks and picnics as shown in the itineraries; • Wines, spirits and Taittinger champagne by the glass plus beers, soft drinks and hot drinks; • Private coach transfers from the local airport or train station to and from Hebridean Princess. For those wishing to drive, parking is provided at the port; • Knowledgeable on board guides; • Renowned guest speakers on selected cruises; • Use of the ship’s bicycles; • Fishing trips and speedboat rides; • Internet access and complimentary Wi-Fi; • All gratuities and port taxes. Hebridean Princess The Ultimate Escape from the Everyday World Whilst travel plans have been placed on hold for many of us recently, most of you will have been dreaming of better times and looking forward to future adventures. The attraction of cruising closer to home, and the appeal of small-ship cruising, make Hebridean Princess the ideal getaway as she sails off the beaten track to remote destinations and secluded anchorages. Every aspect of a holiday on board Hebridean Princess adds to the experience of total relaxation. From the sublime landscapes of the Scottish Highlands and Islands and the spectacular Norwegian fjords, to a genuine welcome, cosseted ambience and impeccable service. -

Hutchesonian 2015
THE HUTCHESONIAN 2015 VERITAS 1 THe HuTcHesonian 2015 conTenTs 2015 Team Editor William Byam Shaw Sub Editors Art and Cover Design Olivia Taylor from THe sTaff noTes communiTy Drama recTor 4 8 23 Melissa Rutnagur Music Elaine Duncan Anna Heywood Sport Katie Turnbull Kingarth Street Hector Ritchie Community/Trips s6 beyonD TriPs anD Amy Greig Creative Writing 30 HuTcHie 35 excHanges 38 Jamie Rodney Photography Professional Photography: JSH Pix, SNS Photography, YoungMedia, Warren Media. Sports Team Photography: cLubs anD Drama anD creaTive © The School socieTies 41 music 48 WriTing 55 Photography Company (www.schoolphotograp hs.co.uk) In-house Photography: Maria Campbell, Kate Flannigan, Anna Heywood (S6), Hutchesons’ Photography Club, Hutchesons’ Form & Year Tutors, KingarTH sPorT Prize LisT Gillian Tooth. Overview sTreeT 63 83 106 Miss Kate Marson Mrs Jane Ritchie Hutchesons’ Grammar School is governed by Hutchesons’ Educational Trust, Registered Charity Number SC002922 2 eDiToriaL o say that 2014 was a big year for Scotland would be a bit of an understatement. The Glasgow TCommonwealth Games was the starting gun that sounded Scotland’s stage-left appearance onto the world stage, with people from lands far and wide flocking to our nation to see what we had to offer, and they weren’t disappointed. For two weeks our country became an international hotspot – the perfect storm of contrasting cultures and integrating nationalities. Our city’s experience during this time can only truly be illustrated through one encounter I had during the Games: I was enjoying a nice walk through a particularly rainy Glasgow Green, before finding myself face-to-face with an athlete from the Malawian team: half- draped in his own country’s flag and clearly plied plenty with some of our country’s good whiskey, he was belting out “Scotland the Brave” at the top of his lungs, whilst also managing to take some selfies with the locals. -

2019 CRUISE DIRECTORY Highlands and Islands of Scotland Orkney and Shetland Northern Ireland and the Isle of Man Cape Wrath Scrabster
Despite the modern fashion for large floating resorts, we b 7 nights 0 2019 CRUISE DIRECTORY Highlands and Islands of Scotland Orkney and Shetland Northern Ireland and The Isle of Man Cape Wrath Scrabster SCOTLAND Kinlochbervie Wick and IRELAND HANDA ISLAND Loch a’ FLANNAN Stornoway Chàirn Bhain ISLES LEWIS Lochinver SUMMER ISLES NORTH ST KILDA Tarbert SHIANT ISLES SEA Ullapool HARRIS Loch Ewe Loch Broom BERNERAY Trotternish Inverewe ATLANTIC NORTH Peninsula OCEAN UIST North Inner Gairloch Minch Sound INVERGORDON Lochmaddy Uig Shieldaig BENBECULA Dunvegan RAASAY INVERNESS SKYE Portree Loch Carron Loch Harport Kyle of Plockton SOUTH Lochalsh UIST Lochboisdale Loch Coruisk Little Minch Loch Hourn ERISKAY CANNA Armadale BARRA RUM Inverie Castlebay Sound of VATERSAY Sleat SCOTLAND PABBAY EIGG MINGULAY MUCK Fort William BARRA HEAD Sea of the Hebrides Glenmore Loch Linnhe Kilchoan Bay Salen COLL CARNA Ballachulish Sound Loch Sunart Tobermory Loch à Choire TIREE of Mull ULVA MULL ISLE OF ERISKA LUNGA Craignure Dunsta!nage STAFFA OBAN IONA KERRERA Firth of Lorn Craobh Haven Inveraray Ardfern Strachur Crarae Loch Goil COLONSAY Crinan Loch Loch Long Tayvallich Rhu LochStriven Fyne JURA Holy Loch GREENOCK Tarbert Portavadie Loch na Mile GLASGOW ISLAY Rothesay BUTE Largs GIGHA GREAT CUMBRAE Port Ellen Lochranza LITTLE CUMBRAE Brodick HOLY Troon ARRAN ISLE Campbeltown Firth of Clyde RATHLIN ISLAND SANDA ISLAND AILSA Ballycastle CRAIG North Channel NORTHERN Larne IRELAND Bangor BELFAST ENGLAND Strangford Lough IRISH SEA ISLE OF MAN EIRE Peel Douglas ORKNEY and Muckle Flugga UNST SHETLAND Baltasound YELL Burravoe Lunna Voe WHALSAY SHETLAND Lerwick Scalloway BRESSAY Grutness FAIR ISLE ATLANTIC OCEAN WESTRAY SANDAY STRONSAY ORKNEY Stromness Kirkwall Scapa Flow HOY Lyness SOUTH RONALDSAY NORTH SEA Pentland Firth STROMA Scrabster Caithness Wick Welcome to the 2019 Hebridean Princess Cruise Directory Unlike most cruise companies, Hebridean operates just one very small and special ship – Hebridean Princess. -

Education in Scotland
EDUCATION IN SCOTLAND Education in Scotland is markedly different from that in the rest of the United Kingdom - with a different curriculum, school boards to oversee school management and a General Teaching Council which has been in existence since 1965. Whilst there are many examples of successful and innovative practice in Scotland, the system is quite often not recognised as different by writers who talk about the United Kingdom education system as if it were one smooth whole. This book describes recent developments in both legislation and practice in Scotland, drawing comparisons with the English system. Chapters cover adminis tration and management, early years education provision, the curriculum in Scotland, secondary education and special educa - tional needs. Margaret Clark is Emeritus Professor of Education at the Uni versity of Birmingham. Pamela Munn is Professor of Curriculum Research at Moray House Institute of Education, Edinburgh. EDUCATION IN SCOTLAND Policy and practice from pre-school to secondary Edited by Margaret M. Clark and Pamela Munn First published 1997 by Routledge Published 2017 by Routledge 2 Park Square, Milton Park, Abingdon, Oxon OX14 4RN 711 Third Avenue, New York, NY 10017, USA Routledge is an imprint ofthe Taylor & Francis Group, an iriforma business Copyright© 1997 Margaret M. Oark, Pamela Munn and individual chapters to their authors The Open Access version of this book, available at www.tandfebooks.com, has been made available under a Creative Commons Attribution-Non Commercial-No Derivatives 4.0 license. British Library Cataloguing in Publication Data A catalogue record for this book is available from the British Library Library of Congress Cataloguing in Publication Data Education in Scotland: policy and practice from pre-school to secondary/edited by Margaret M. -

Seabird Numbers and Breeding Success in Britain and Ireland, 2004
UK Nature Conservation No. 29 Seabird numbers and breeding success in Britain and Ireland, 2004 R.A. Mavor, M. Parsons, M. Heubeck and S. Schmitt Roddy Mavor and Matt Parsons, Seabirds and Cetaceans, Joint Nature Conservation Committee, Dunnet House, 7 Thistle Place, Aberdeen, AB10 1UZ Martin Heubeck, Shetland Oil Terminal Environmental Advisory Group, Department of Zoology, University of Aberdeen, Tillydrone Avenue, Aberdeen, AB9 2TN Sabine Schmitt, Research Department, Royal Society for the Protection of Birds, The Lodge, Sandy, Bedfordshire, SG19 2DL Cover painting of black-legged kittiwakes by David Bennett. Cover design by Green Associates, 1994. Further information on JNCC publications can be obtained from: JNCC, Monkstone House, City Road, Peterborough PE1 1JY Published by Joint Nature Conservation Committee, Peterborough Copyright: Joint Nature Conservation Committee 2005 ISBN 1 86107 569 3 ISSN 0963 8083 Seabird numbers and breeding success, 2004 This report should be cited as follows: Mavor, R.A., Parsons, M., Heubeck, M. and Schmitt, S. 2005. Seabird numbers and breeding success in Britain and Ireland, 2004. Peterborough, Joint Nature Conservation Committee. (UK Nature Conservation, No. 29.) Original data from the report may not be used in other publications (although general results and conclusions may be cited) without permission of JNCC, RSPB and/or SOTEAG, as relevant. Requests to use data should be channelled through S. Schmitt/RSPB (petrels, terns and skuas), M. Heubeck/SOTEAG (cliff-breeding species and black guillemots