Why Organizational Ecology Is Not a Darwinian Research Program
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Degrees, Academic Offerings, and Accreditations
Degrees, Academic Offerings, and Accreditations Associate degrees A.A. -- Associate of Arts A.A.S. -- Associate of Applied Science A.S. -- Associate of Science A.S.N. -- Associate of Science in Nursing Bachelor’s degrees B.A.-- Bachelor of Arts B.F.A.-- Bachelor of Fine Arts B.M..-- Bachelor of Music B.S.-- Bachelor of Science B.S.B.A.-- Bachelor of Science in Business Administration B.S.Ed.-- Bachelor of Science in Education B.S.N.-- Bachelor of Science in Nursing Master’s degrees M.A.-- Master of Arts M.B.A.-- Master of Business Administration M.Ed.-- Master of Education M.S.-- Master of Science M.S.L.S.-- Master of Science in Library Science M.S.N.-- Master of Science in Nursing Academic Offerings at Clarion University *Academic Concentrations Bachelor and Master’s Degrees Accounting (BSBA) Anthropology (BA) Art (BA, BFA) Ceramics* Drawing* Dual Drawing* Fiber/Fabrics* Graphic Arts* Painting* Printmaking* Sculpture* Athletic Training (BSAT) from California University of Pennsylvania Biology (MS) Biological Sciences* (graduate levels) Environmental Sciences* (graduate levels) Biology (BA, BS) Ecology and Evolutionary Biology* Business Administration (MBA) Accountancy* Economics* Finance* International Business* Management* Marketing* Chemistry (BS) Biochemistry* Chemistry/Business* Chemistry/Cooperative Engineering* Computer Science (BS) Early Childhood Education (BSED) Economics (BA) Economics, Business (BSBA) Education (MED) Coaching* Curriculum and Instruction* Early Childhood* English* History* Literacy* Mathematics* Reading Specialist* -

Business Studies Stage 6 Syllabus
Business Studies Stage 6 Syllabus June 2010 Original published version updated: August 2013 – Updated with minor amendments © 2010 Copyright Board of Studies NSW for and on behalf of the Crown in right of the State of New South Wales. This document contains Material prepared by the Board of Studies NSW for and on behalf of the State of New South Wales. The Material is protected by Crown copyright. All rights reserved. No part of the Material may be reproduced in Australia or in any other country by any process, electronic or otherwise, in any material form or transmitted to any other person or stored electronically in any form without the prior written permission of the Board of Studies NSW, except as permitted by the Copyright Act 1968. School students in NSW and teachers in schools in NSW may copy reasonable portions of the Material for the purposes of bona fide research or study. When you access the Material you agree: • to use the Material for information purposes only • to reproduce a single copy for personal bona fide study use only and not to reproduce any major extract or the entire Material without the prior permission of the Board of Studies NSW • to acknowledge that the Material is provided by the Board of Studies NSW • not to make any charge for providing the Material or any part of the Material to another person or in any way make commercial use of the Material without the prior written consent of the Board of Studies NSW and payment of the appropriate copyright fee • to include this copyright notice in any copy made • not to modify the Material or any part of the material without the express prior written permission of the Board of Studies NSW. -

New Renovated B-School Facilities
Business School Facilities: Recent Construction and Renovation Institution Name B-school Name Building/Facility Name Activity Year Status University of Calgary Haskayne School of Business Scurfield Hall New Building 1986 Complete University of Cincinnati School of Business Carl H. Lindner Hall New Building 1987 Complete Brock University Faculty of Business Taro Hall New Building 1990 Complete The University of Arizona Eller College of Management McClelland Hall New Building 1992 Complete University of California, Berkeley Haas School of Business Haas School of Business complex New Building 1995 Complete University of California, Los Angeles Anderson School of Management Management Education Complex New Building 1995 Complete Boston University School of Management Rafik B. Hariri Building New Building 1996 Complete Creighton University College of Business College of Business Building Renovation/Expansion 1996 Complete Northern Kentucky University Haile/US Bank College of Business unknown unknown 1996 Complete University of Georgia The Terry College of Business Brooks Hall Renovation/Expansion 1996 Complete William and Rosemary Gallagher University of Montana School of Business Administration Business Building New Building 1996 Complete University of Virginia-Darden Darden Graduate School of Business Saunders Hall New Building 1996 Complete The Arnold and Mabel Beckman Chapman University Argyros School of Business and Economics Business and Technology Hall New Building 1997 Complete Peter F. Drucker & Masatoshi Ito Graduate Claremont Graduate -

B.S. Business Studies Management Concentration
All Bachelor of Science programs require a minimum of 128 credits and a 2.00 cumulative average. B.S. BUSINESS STUDIES MANAGEMENT CONCENTRATION Fall 2015 – Spring 2016 BSNS REQUIREMENTS: All BSNS program courses must be completed with a grade of “C” or 80 credits better. CSIS 1206 Statistics (4) FINA 3562 Budgeting (4) ECON 1200 Macroeconomics (4) MGMT 3111 Human Resource Mgmt. (4) ECON 1400 Microeconomics (4) MGMT 3121 Mgmt. Info. Systems (4) ACCT 2110 Financial ACCT (4) MGMT 3123 Mgmt. Skills Development (4) ACCT 2120 Managerial ACCT (4) MGMT 3124 Organizational Behavior (4) BSNS 2120 Quantitative BSNS Methods (4) MGMT Elective (4) MGMT 2110 Intro to Management (4) MGMT 4610 Senior Seminar OR (4) MKTG 2110 Marketing Principles (4) MGMT Elective PLAW 2120 Business Law I OR (4) Internship or BSNS Elective: Choose (4) PLAW 3110 Legal, Social, Ethical … from ACCT, ECON, FINA, HTMS, INTL, FINA 3110 Intro to Financial Mgmt* (4) MKTG, MGMT, PLAW at the 3000 level MGMT 3120 Operations Management (4) or above BSNS 4112 Business Policy & Strategies (4) Transfer students may use transferred courses (seniors only) (including Introduction to Business) as “Other Business Courses”, to satisfy the minimum number of credits (80) for this area. GENERAL STUDIES REQUIREMENTS: 48 credits G COURSES: (32 total credits) No more than 12 credits in any "G" category may be applied towards the BS degree. GEN General Interdisciplinary (4) GNM General Natural Science & Math (4) GIS-General Integration & Synthesis (Jr. yr.) (4) GNM General Natural Science & Math (4) GAH General Arts & Humanities (4) GSS General Social Science (4) GAH General Arts & Humanities (4) GSS General Social Science (4) AT SOME DISTANCE Electives: (16 total credits) Courses unrelated to your major (may include CSIS courses) (4) (4) (4) (4) GENERAL STUDIES OUTCOME REQUIREMENTS: These course attributes should be completed within the 128 credits needed to graduate. -

Business Studies
Programs BUS.STUD.AAS Business Studies BUS.STUD.MGMT.AAS BUS.STUD.ENTR.AAS Associate in Applied Science Degree in Business Management CIP 520101 Business Studies, a career and non-transferable The Management concentration (BUS.STUD.MGMT. degree program, provides opportunities for students AAS) prepares students for positions as general office to prepare for a wide variety of careers in business. managers, retail sales managers, or supervisors in mid- Awarded an A.A.S. degree in Business Management, to large-sized firms. Students focus their degree based successful graduates of the program will be able to: on individual professional needs. In addition to the com- • use effective verbal and written communication petencies gained from the general Business Studies pro- in conducting business; gram, students completing this concentration will be able • analyze/resolve problems common to entry-level to (depending on electives chosen): business positions; • understand the laws affecting the domestic business • apply management skills in a variety of business environment; functions; • apply the basic principles of marketing to business; • comprehend how the global economy and • apply a better understanding of the customer’s Business Studies international events affect domestic and behavior in improving sales management; international business decisions; • lead and/or manage an organization’s human • understand basic accounting statements and resources on both an individual and team level their role in managing a business; to maximize workforce effectiveness. • identify unethical behavior in a business setting and formulate appropriate action. The Entrepreneurship concentration (BUS.STUD. Students have the option of concentrating in Man- ENTR.AAS) prepares students to own a small business. -

Talent Management Impacting Performance
Using Action Research to Support an On-going Organizational Culture Transformation in a Health Care Organization A Dissertation SUBMITTED TO THE FACULTY OF THE UNIVERSITY OF MINNESOTA BY Jane Marie Kuhn IN PARTIAL FULFILLMENT OF THE REQUIREMENTS FOR THE DEGREE OF DOCTOR OF PHILOSOPHY Co-advisers Dr. Deanne Magnusson and Dr. Louis N. Quast September 2016 © Jane Kuhn 2016 Acknowledgments This research study would not have been possible without the support, participation, and guidance of many people. Thanks to my organization, which supported me in my endeavor to advance my education, combined with allowing internal research and encouraging its implementation. Without the organization’s leadership support, this study would not have been possible. Thank you for the contributions of the CART team members who participated in this research study. They willingly participated in the action research process, including reflective practices to determine lessons learned. They created a tight-knit community driven by the unwavering commitment to improve the organization, while developing relationships. I appreciated their openness and enthusiasm, making it a much richer experience than anticipated. In addition to these team members, many organizational experts supported the project and this research study. Thanks to Internal Communications, the Creative Marketing Team, Talent Recruiting, and the Talent Analytics team. Their expertise and commitment helped drive to completion with creativity and a great end product. Thanks to my co-advisers, Dr. Deanne Magnusson, Dr. Shari Peterson, and Dr. Louis N. Quast, who continued to support me throughout the years, as we discussed many research options and encouraged me to complete the research. Many thanks to my partner, Rick Benesh, for his help and support during this educational journey. -
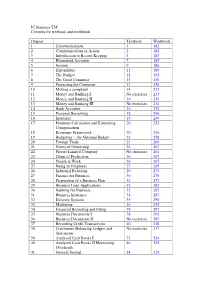
Junior Certificate Business Studies for Households and Enterprises TEXTBOOK TEACHER’S MANUAL
JC Business TM Contents for textbook and workbook Chapter Textbook Workbook 1 Communications 1 182 2 Communications in Action 2 182 3 Introduction to Record Keeping 5 183 4 Household Accounts 7 185 5 Income 9 186 6 Expenditure 11 189 7 The Budget 12 192 8 The Good Consumer 13 230 9 Protecting the Consumer 13 230 10 Making a complaint 14 232 11 Money and Banking I No exercises 233 12 Money and Banking II 16 234 13 Money and Banking III No exercises 234 14 Bank Accounts 16 238 15 Personal Borrowing 18 246 16 Insurance 19 249 17 Premium Calculation and Estimating 20 252 Compensation 18 Economic Framework 20 256 19 Budgeting - the National Budget 21 258 20 Foreign Trade 23 260 21 Forms of Ownership 24 263 22 Private Limited Company No exercises 264 23 Chain of Production 26 267 24 People at Work 26 267 25 Being an Employer 27 269 26 Industrial Relations 29 275 27 Finance for Business 30 276 28 Preparation of a Business Plan 32 277 29 Business Loan Applications 32 283 30 Banking for Business 32 287 31 Business Insurance 34 287 32 Delivery Systems 35 290 33 Marketing 36 295 34 Financial Recording and Filing 38 297 35 Business Documents I 38 302 36 Business Documents II No exercises 305 37 Recording Credit Transactions 40 310 38 Continuous Balancing Ledgers and No exercises 317 Statements 39 Analysed Cash Books I 71 324 40 Analysed Cash Books II Monitoring 80 325 Overheads 41 General Journal 84 329 42 Combined Books of First Entry and 91 331 Ledger Questions 43 Control Accounts 109 333 44 Trading Account 109 334 45 Profit and Loss Account 111 -

Glenn R. Carroll Source: Annual Review of Sociology, Vol. 10 (1984), Pp
Organizational Ecology Author(s): Glenn R. Carroll Source: Annual Review of Sociology, Vol. 10 (1984), pp. 71-93 Published by: Annual Reviews Stable URL: http://www.jstor.org/stable/2083168 Accessed: 07-03-2016 21:52 UTC Your use of the JSTOR archive indicates your acceptance of the Terms & Conditions of Use, available at http://www.jstor.org/page/ info/about/policies/terms.jsp JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range of content in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new forms of scholarship. For more information about JSTOR, please contact [email protected]. Annual Reviews is collaborating with JSTOR to digitize, preserve and extend access to Annual Review of Sociology. http://www.jstor.org This content downloaded from 143.107.252.192 on Mon, 07 Mar 2016 21:52:02 UTC All use subject to JSTOR Terms and Conditions Ann. Rev. Sociol. 1984. 10:71-93 Copyright ? 1984 by Annual Reviews Inc. All rights reserved ORGANIZATIONAL ECOLOGY Glenn R. Carroll School of Business Administration, University of California, Berkeley, California 94720 Abstract Recent research on organizational ecology is reviewed. Three levels of analysis and approaches to evolution are distinguished: (a) the organizational level, which uses a developmental approach; (b) the population level, which uses a selection approach; and (c) the community level, which uses a macroevolution- ary approach. Theoretical and empirical research is critiqued within this framework. Proposals to develop organizational taxonomies are considered. INTRODUCTION Although the intellectual descendant of human ecology (e.g. -

Department of Business Studies
Department of Business Studies Associate Professors: Joseph T. Foley (Chairperson), Daniel J. Jones, Francis A. Marino, J. Bart Morrison, Jennifer M. Niece (Coordinator); Assistant Professors: Arlene DeWitt, Robin Frkal, Cary LeBlanc, Bridget Leonard, Michael Lewis, Elizabeth O’Hara, Catherine L. Pastille; Professors of Practice: Eric Drouart, Megan Hill, Kevin Kelly; Visiting Assistant Professors: Paul Bailey, Bryan Coleman, Zachary Daniels, Paul Piwko; Lecturers: Philip Benvenuti, Thomas L. Fitzpatrick, Mary Kingsley, Justin Lundberg, Michael Matraia. The Department of Business Studies offers majors and minors that, when combined with substantial background in the liberal arts disciplines, provide our students with the knowledge and skills valued by the business world. Our majors also focus on helping students build socially responsible lives with ethical cores. MISSION STATEMENT The Business Studies Department of Assumption College offers undergraduate programs of study in accounting, international business, management, marketing, and organizational communication and a graduate business program with concentrations in management, marketing, accounting, finance, and international business. These programs are part of the College’s professional studies. The general mission of the Business Studies Department is to create a challenging learning environment that prepares students for professional employment in the private, public, and nonprofit sectors and/or graduate study. Assumption College has a strong liberal arts tradition, and Business -

Economics and Business Studies - Curriculum Intent, Implementation and Impact
NHGS - Economics and Business Studies - Curriculum Intent, Implementation and Impact Intent (Aims and purpose) We aim to create the very best Economics and Business studies students. The aim of the Economics curriculum is to develop students’ understanding of how the local / national / global economy works through analysing economic issues, problems and institutions that affect everyday life.The aim of the Business studies curriculum is to equip students with the appropriate knowledge and skills needed to develop their employability and identify business problems and opportunities. We do this by quality first teaching which ensures students understand underlying Economic and Business theory which students apply to a variety of familiar and unfamiliar case studies. We want students to be able to think analytically, reach logical conclusions based on data, and make judgements on future changes to markets and the economy. Our curriculum at NHGS goes far beyond what is taught in lessons, for whilst we want students to achieve the very best examination results possible, we believe our curriculum goes beyond what is examinable. Our curriculum in Economics and Business studies supports the ethos statement of the school. Students are constantly challenged to work collaboratively and think independently when engaging in all lessons and class debates. Through teacher modelling, we encourage our students to demonstrate manners, respect and tolerance in Economics and Business studies lessons. This allows students to express themselves in a confident manner. Lesson materials are engaging to promote topical discussion and encourage students to develop an enquiring mind. As a knowledge based curriculum we believe that knowledge underpins and enables the application of skills; both are entwined. -

Business Studies and Employability Skills Development in Junior Secondary Schools in Ilorin Metropolis, Kwara State Dr
Business Studies and Employability Skills Development in Junior Secondary Schools in Ilorin Metropolis, Kwara State Dr. Felicia Kikelomo Oluwalola Department of Educational Management Faculty of Education University of Ilorin, Nigeria [email protected] Fall 2019 Abstract The study examined business studies and employability skills development in junior secondary schools in Ilorin metropolis Kwara State. Five research questions and three hypotheses guided the study. A survey research designed was adopted for the study. The population of this study comprises all the 4570 teachers in all the private and public junior secondary schools and the employers in Ilorin Metropolis, Kwara State. However the target population includes all the 1305 business studies related teachers in all the junior secondary schools in Ilorin Metropolis, Kwara State. The sample of the study includes 250 teachers in all the junior secondary schools and 250 employers in Ilorin Metropolis, Kwara State. A structured questionnaire titled “Business Studies Questionnaire” (BSQ) and “Employability Skill Development” (ESDQ) were used to obtain relevant data. Descriptive statistics of mean and standard deviation while inferential statistics of Pearson Product-Moment correlation was used to test the hypotheses formulated at 0.05 level of significance. The findings revealed that level of knowledge acquisition in business studies was rated high by business studies teachers, employability skills level of junior secondary schools students were rated high by the employers of labour also, there was positive relationship exists between accounting, office practice, marketing and employability skills development in junior secondary school Ilorin Metropolis Kwara State. The study concluded that the students that graduated within Ilorin metropolis are possessed with basic soft skills; digital, technical and analytical skills required that could make them employable after graduation. -

Business Studies Grade 12 Term Two Chapter 6 Notes Csr and Csi 2019
CSR AND CSI CHAPTER 6 BUSINESS STUDIES GRADE 12 TERM TWO CHAPTER 6 NOTES CSR AND CSI 2019 TABLE OF CONTENTS TOPICS PAGES Exam guidelines for CSR and CSI 2 Terms and definitions 3 Meaning of social responsibility 4 Relationship between triple bottom line and 4 social responsibility Socio-economic issues 5 Strategies to deal with HIV/Aids, 5 unemployment and poverty Ways in which businesses could contribute to 5 the wellbeing of their employees and community Meaning of CSR 6 Purpose of CSR 6 Components of Corporate Social 7 Responsibility Impact of CSR/CSI on businesses 7-9 &communities The meaning of CSI 9 Purpose of CSI 9 CSI Focus areas 9 Examples of CSI projects 10 Differences between CSR and CSI 10 1 CSR AND CSI CHAPTER 6 CONTENT DETAILS FOR TEACHING, LEARNING AND ASSESSMENT PURPOSES Learners must be able to: SOCIAL RESPONSIBILITY • Define social responsibility • Explain the relationship/link between social responsibility and triple bottom line. • Suggest ways in which a business project can contribute towards the community, e.g. charitable contributions to NGO's, involvement in community education, employment, employee volunteer programmes, etc. • Identify socio-economic issues that challenge social responsibility, e.g. HIV/Aids, unemployment, poverty, human rights aspects etc. • Recommend/Suggest ways in which businesses can deal with the above mentioned socio-economic issues. • Explain how businesses can contribute time and effort in improving the well-being of the following stakeholders: o Employees o Community COOPERATE SOCIAL RESPONSIBILITY/CSR • Define/Elaborate on the meaning of CSR • Describe/Explain the nature/ purpose of CSR • Identify/Name/Outline the components of CSR, e.g.