Ephemeral Wetlands of the Nelson Mandela Bay Metropolitan Area: Classification, Biodiversity and Management Implications
Total Page:16
File Type:pdf, Size:1020Kb

Load more
Recommended publications
-
Fig. Ap. 2.1. Denton Tending His Fairy Shrimp Collection
Fig. Ap. 2.1. Denton tending his fairy shrimp collection. 176 Appendix 1 Hatching and Rearing Back in the bowels of this book we noted that However, salts may leach from soils to ultimately if one takes dry soil samples from a pool basin, make the water salty, a situation which commonly preferably at its deepest point, one can then "just turns off hatching. Tap water is usually unsatis- add water and stir". In a day or two nauplii ap- factory, either because it has high TDS, or because pear if their cysts are present. O.K., so they won't it contains chlorine or chloramine, disinfectants always appear, but you get the idea. which may inhibit hatching or kill emerging If your desire is to hatch and rear fairy nauplii. shrimps the hi-tech way, you should get some As you have read time and again in Chapter 5, guidance from Brendonck et al. (1990) and temperature is an important environmental cue for Maeda-Martinez et al. (1995c). If you merely coaxing larvae from their dormant state. You can want to see what an anostracan is like, buy some guess what temperatures might need to be ap- Artemia cysts at the local aquarium shop and fol- proximated given the sample's origin. Try incu- low directions on the container. Should you wish bation at about 3-5°C if it came from the moun- to find out what's in your favorite pool, or gather tains or high desert. If from California grass- together sufficient animals for a study of behavior lands, 10° is a good level at which to start. -

Phylogenetic Analysis of Anostracans (Branchiopoda: Anostraca) Inferred from Nuclear 18S Ribosomal DNA (18S Rdna) Sequences
MOLECULAR PHYLOGENETICS AND EVOLUTION Molecular Phylogenetics and Evolution 25 (2002) 535–544 www.academicpress.com Phylogenetic analysis of anostracans (Branchiopoda: Anostraca) inferred from nuclear 18S ribosomal DNA (18S rDNA) sequences Peter H.H. Weekers,a,* Gopal Murugan,a,1 Jacques R. Vanfleteren,a Denton Belk,b and Henri J. Dumonta a Department of Biology, Ghent University, Ledeganckstraat 35, B-9000 Ghent, Belgium b Biology Department, Our Lady of the Lake University of San Antonio, San Antonio, TX 78207, USA Received 20 February 2001; received in revised form 18 June 2002 Abstract The nuclear small subunit ribosomal DNA (18S rDNA) of 27 anostracans (Branchiopoda: Anostraca) belonging to 14 genera and eight out of nine traditionally recognized families has been sequenced and used for phylogenetic analysis. The 18S rDNA phylogeny shows that the anostracans are monophyletic. The taxa under examination form two clades of subordinal level and eight clades of family level. Two families the Polyartemiidae and Linderiellidae are suppressed and merged with the Chirocephalidae, of which together they form a subfamily. In contrast, the Parartemiinae are removed from the Branchipodidae, raised to family level (Parartemiidae) and cluster as a sister group to the Artemiidae in a clade defined here as the Artemiina (new suborder). A number of morphological traits support this new suborder. The Branchipodidae are separated into two families, the Branchipodidae and Ta- nymastigidae (new family). The relationship between Dendrocephalus and Thamnocephalus requires further study and needs the addition of Branchinella sequences to decide whether the Thamnocephalidae are monophyletic. Surprisingly, Polyartemiella hazeni and Polyartemia forcipata (‘‘Family’’ Polyartemiidae), with 17 and 19 thoracic segments and pairs of trunk limb as opposed to all other anostracans with only 11 pairs, do not cluster but are separated by Linderiella santarosae (‘‘Family’’ Linderiellidae), which has 11 pairs of trunk limbs. -

Site Development Plans Standard Operating Procedures for COVID
NELSON MANDELA BAY METROPOLITAN MUNICIPALITY NOTICE SITE DEVELOPMENT PLAN APPLICATIONS (SDP) STANDARD OPERATING PROCEDURES FOR DURING COVID-19 PERIOD A. SITE DEVELOPMENT PLANS (SDPs) 1. SDP applications are to be converted into PDF format PDF (not exceeding 2mb) and submitted per email to the Planning Support Officer Ms N Ketelo at: [email protected], who will in turn acknowledge receipt of the application per email. 2. In the event that the attachments combined exceeds the prescribed size, the application must be split into separate emails or put in a flash-drive to be delivered to Land Planning Division in 3rd Floor Lillian Diedericks Building, 191 Govan Mbeki Avenue, Port Elizabeth, preferably on appointment. 3. Documents must be labelled Annexure A, B, C…etc. as guided by the table appended hereto showing documents required. 4. An assessment sheet will be emailed back to an applicant by a Town Planner dealing with the SDP. 5. It is the responsibility of an applicant to obtain comments from various municipal departments. 6. An applicant must make amendments to the plan as per comments received from various municipal departments, if required. 7. An applicant must submit the plan and the assessment sheet in PDF (email or flash-drive) to a Town Planner dealing with SDP through email. 8. A Town Planner will acknowledge receipt, finalize the SDP and communicate the outcome to an applicant through a correspondence sent via email. B. TOWN PLANNING COMMENTS ON BUILDING PLANS 1. Technical Controllers responsible for Town Planning related comments made on building plans will be available on Thursdays for inquiries at 3rd Floor Lillian Diedericks Building, Govan Mbeki Avenue, Port Elizabeth. -

Position Specificity in the Genus Coreomyces (Laboulbeniomycetes, Ascomycota)
VOLUME 1 JUNE 2018 Fungal Systematics and Evolution PAGES 217–228 doi.org/10.3114/fuse.2018.01.09 Position specificity in the genus Coreomyces (Laboulbeniomycetes, Ascomycota) H. Sundberg1*, Å. Kruys2, J. Bergsten3, S. Ekman2 1Systematic Biology, Department of Organismal Biology, Evolutionary Biology Centre, Uppsala University, Uppsala, Sweden 2Museum of Evolution, Uppsala University, Uppsala, Sweden 3Department of Zoology, Swedish Museum of Natural History, Stockholm, Sweden *Corresponding author: [email protected] Key words: Abstract: To study position specificity in the insect-parasitic fungal genus Coreomyces (Laboulbeniaceae, Laboulbeniales), Corixidae we sampled corixid hosts (Corixidae, Heteroptera) in southern Scandinavia. We detected Coreomyces thalli in five different DNA positions on the hosts. Thalli from the various positions grouped in four distinct clusters in the resulting gene trees, distinctly Fungi so in the ITS and LSU of the nuclear ribosomal DNA, less so in the SSU of the nuclear ribosomal DNA and the mitochondrial host-specificity ribosomal DNA. Thalli from the left side of abdomen grouped in a single cluster, and so did thalli from the ventral right side. insect Thalli in the mid-ventral position turned out to be a mix of three clades, while thalli growing dorsally grouped with thalli from phylogeny the left and right abdominal clades. The mid-ventral and dorsal positions were found in male hosts only. The position on the left hemelytron was shared by members from two sister clades. Statistical analyses demonstrate a significant positive correlation between clade and position on the host, but also a weak correlation between host sex and clade membership. These results indicate that sex-of-host specificity may be a non-existent extreme in a continuum, where instead weak preferences for one host sex may turn out to be frequent. -

Table of Contents 2
Southwest Association of Freshwater Invertebrate Taxonomists (SAFIT) List of Freshwater Macroinvertebrate Taxa from California and Adjacent States including Standard Taxonomic Effort Levels 1 March 2011 Austin Brady Richards and D. Christopher Rogers Table of Contents 2 1.0 Introduction 4 1.1 Acknowledgments 5 2.0 Standard Taxonomic Effort 5 2.1 Rules for Developing a Standard Taxonomic Effort Document 5 2.2 Changes from the Previous Version 6 2.3 The SAFIT Standard Taxonomic List 6 3.0 Methods and Materials 7 3.1 Habitat information 7 3.2 Geographic Scope 7 3.3 Abbreviations used in the STE List 8 3.4 Life Stage Terminology 8 4.0 Rare, Threatened and Endangered Species 8 5.0 Literature Cited 9 Appendix I. The SAFIT Standard Taxonomic Effort List 10 Phylum Silicea 11 Phylum Cnidaria 12 Phylum Platyhelminthes 14 Phylum Nemertea 15 Phylum Nemata 16 Phylum Nematomorpha 17 Phylum Entoprocta 18 Phylum Ectoprocta 19 Phylum Mollusca 20 Phylum Annelida 32 Class Hirudinea Class Branchiobdella Class Polychaeta Class Oligochaeta Phylum Arthropoda Subphylum Chelicerata, Subclass Acari 35 Subphylum Crustacea 47 Subphylum Hexapoda Class Collembola 69 Class Insecta Order Ephemeroptera 71 Order Odonata 95 Order Plecoptera 112 Order Hemiptera 126 Order Megaloptera 139 Order Neuroptera 141 Order Trichoptera 143 Order Lepidoptera 165 2 Order Coleoptera 167 Order Diptera 219 3 1.0 Introduction The Southwest Association of Freshwater Invertebrate Taxonomists (SAFIT) is charged through its charter to develop standardized levels for the taxonomic identification of aquatic macroinvertebrates in support of bioassessment. This document defines the standard levels of taxonomic effort (STE) for bioassessment data compatible with the Surface Water Ambient Monitoring Program (SWAMP) bioassessment protocols (Ode, 2007) or similar procedures. -

Luc Strydom Environmental Consultant
SRK Consulting Page 1 Luc Strydom Environmental Consultant Profession Environmental Scientist Education BA Environmental Management, University of South Africa, 2015 Registrations/ Registered EAP, EAPASA (2020/1504) Affiliations Certificated Natural Scientist (EIA), SACNASP (Reg No. 120385) Member, South African Wetland Society (Membership No.: 193665) Member, International Association of Impact Assessors, South Africa (IAIAsa), Volunteer, Custodians of Rare and Endangered Wildflowers (CREW). Specialisation Wetland and aquatic impact assessments, botanical surveys, vegetation impact assessments, invasive alien monitoring and control plans, rehabilitation plans, environmental impact and basic assessments, environmental management programmes (EMPrs), water use license applications (WULAs), environmental auditing (environmental control officer), geo-hydrological sampling, section 24G applications & GIS systems. Expertise Luc Strydom has previous experience in GIS, working for Setplan PE, a town planning consultancy group. His expertise in GIS includes map production, data capturing, data manipulation, data acquisition and database management. Luc has developed his skills and expertise over the years as he has been involved in many different types of environmental projects, such as: • environmental impact assessments (EIAs); • wetland and aquatic impact assessments (wetland screening, delineation, PES & EIS determination, ecosystem services assessment, etc.); • environmental management plans/programmes (EMPr); • environmental auditing (acting -
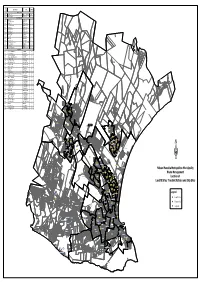
Map of Landfill Transfer and Skip Sites
Origin DESCRIPTION AREA WARD Code LANDFILL SITES G01 ARLINGTON PORT ELIZABETH 01 G02 KOEDOESKLOOF UITENHAGE 53 TRANSFER SITES G03 TAMBO STR McNAUGHTON 50 G04 GILLESPIE STR EXT UITENHAGE 53 G05 JOLOBE STR KWANOBUHLE 47 G06 ZOLILE NOGCAZI RD KWANOBUHLE 45 G07 SARILI STR KWANOBUHLE 42 G08 VERWOERD RD UITENHAGE 51 G09 NGEYAKHE STR KWANOBUHLE 42 G10 NTAMBANANI STR KWANOBUHLE 47 G11 LAKSMAN AVE ROSEDALE 49 G12 RALO STR KWAMAGXAKI 30 G13 GAIL ROAD GELVANDALE 13 G14 DITCHLING STR ALGOA PARK 11 G15 CAPE ROAD - OPP HOTEL HUNTERS RETREAT 12 G16 KRAGGAKAMA RD FRAMESBY 09 G17 5TH AVENUE WALMER 04 G18 STRANDFONTEIN RD SUMMERSTRAND 02 SKIP SITES G19 OFF WILLIAM MOFFAT FAIRVIEW 06 G20 STANFORD ROAD HELENVALE 13 G21 OLD UITENHAGE ROAD GOVAN MBEKI 33 G22 KATOO & HARRINGTON STR SALTLAKE 32 G23 BRAMLIN STREET MALABAR EXT 6 09 G24 ROHLILALA VILLAGE MISSIONVALE 32 G25 EMPILWENI HOSPITAL NEW BRIGHTON 31 G26 EMLOTHENI STR NEW BRIGHTON 14 G27 GUNGULUZA STR NEW BRIGHTON 16 G28 DAKU SQUARE KWAZAKHELE 24 G29 DAKU SPAR KWAZAKHELE 21 G30 TIPPERS CREEK BLUEWATER BAY 60 G31 CARDEN STR - EAST OF RAILWAY CROSSING REDHOUSE 60 G32 BHOBHOYI STR NU 8 MOTHERWELL 56 G33 UMNULU 1 STR NU 9 MOTHERWELL 58 Farms Uitenhage G34 NGXOTWANE STR NU 9 MOTHERWELL 57 G51 ward 53 S G35 MPONGO STR NU 7 MOTHERWELL 57 G36 MGWALANA STR NU 6 MOTHERWELL 59 G37 DABADABA STR NU 5 MOTHERWELL 59 G38 MBOKWANE NU 10 MOTHERWELL 55 Colchester G39 GOGO STR NU 1 MOTHERWELL 56 G40 MALINGA STR WELLS ESTATE 60 G41 SAKWATSHA STR NU 8 MOTHERWELL 58 G42 NGQOKWENI STR NU 9 MOTHERWELL 57 G43 UMNULU 2 STR NU 9 MOTHERWELL -

Aia) for the Proposed Mixed-Use Housing Development, Kwanobuhle, Extension 11, Uitenhage, Nelson Mandela Bay Muncipality, Eastern Cape Province
A PHASE 1 ARCHAEOLOGICAL IMAPCT ASSESMENT (AIA) FOR THE PROPOSED MIXED-USE HOUSING DEVELOPMENT, KWANOBUHLE, EXTENSION 11, UITENHAGE, NELSON MANDELA BAY MUNCIPALITY, EASTERN CAPE PROVINCE Prepared for: SRK Consulting PO Box 21842 Port Elizabeth 6000 Tel: 041 509 4800 Fax: 041 509 4850 Contact person: Ms Karissa Nel Email: [email protected] Compiled by: Dr Johan Binneman, Ms Celeste Booth and Ms Natasha Higgitt Department of Archaeology Albany Museum Somerset Street Grahamstown 6139 Tel: (046) 622 2312 Fax: (046) 622 2398 Contact person: Ms. Celeste Booth Email: [email protected] July 2011 1 TABLE OF CONTENTS EXECUTIVE SUMMARY 2. BACKGROUND INFORMATION 3. BRIEF ARCHAEOLOGICAL BACKGROUND 6. DESCRIPTION OF THE PROPERTY 7. ARCHAEOLOGICAL INVESTIGATION 8. CULTURAL LANDSCAPE 11. RECOMMENDATIONS 12. GENERAL REMARKS AND CONDITIONS 13. APPENDIX A 14. MAP 1 16. MAP 2 17. MAP 3 18. TABLE 1 19. 2 A PHASE 1 ARCHAEOLOGICAL IMAPCT ASSESMENT (AIA) FOR THE PROPOSED MIXED-USE HOUSING DEVELOPMENT, KWANOBUHLE, EXTENSION 11, UITENHAGE, NELSON MANDELA BAY MUNCIPALITY, EASTERN CAPE PROVINCE Note: This report follows the minimum standard guidelines required by the South African Heritage Resources Agency for compiling a Phase 1 Archaeological Impact Assessment (AIA). EXECUTIVE SUMMARY Purpose of the Study The purpose of the study was to conduct a phase 1 archaeological impact assessment (AIA) for the proposed mixed-use housing development, Kwanobuhle Extension 11, Uitenhage, Nelson Mandela Bay Municipality, Eastern Cape Province. The survey was conducted to establish the range and importance of the exposed and in situ archaeological heritage materials and features, the potential impact of the development, and to make recommendations to minimize possible damage to these sites. -

This Table Contains a Taxonomic List of Benthic Invertebrates Collected from Streams in the Upper Mississippi River Basin Study
This table contains a taxonomic list of benthic invertebrates collected from streams in the Upper Mississippi River Basin study unit as part of the USGS National Water Quality Assessemnt (NAWQA) Program. Invertebrates were collected from woody snags in selected streams from 1996-2004. Data Retreival occurred 26-JAN-06 11.10.25 AM from the USGS data warehouse (Taxonomic List Invert http://water.usgs.gov/nawqa/data). The data warehouse currently contains invertebrate data through 09/30/2002. Invertebrate taxa can include provisional and conditional identifications. For more information about invertebrate sample processing and taxonomic standards see, "Methods of analysis by the U.S. Geological Survey National Water Quality Laboratory -- Processing, taxonomy, and quality control of benthic macroinvertebrate samples", at << http://nwql.usgs.gov/Public/pubs/OFR00-212.html >>. Data Retrieval Precaution: Extreme caution must be exercised when comparing taxonomic lists generated using different search criteria. This is because the number of samples represented by each taxa list will vary depending on the geographic criteria selected for the retrievals. In addition, species lists retrieved at different times using the same criteria may differ because: (1) the taxonomic nomenclature (names) were updated, and/or (2) new samples containing new taxa may Phylum Class Order Family Subfamily Tribe Genus Species Taxon Porifera Porifera Cnidaria Hydrozoa Hydroida Hydridae Hydridae Cnidaria Hydrozoa Hydroida Hydridae Hydra Hydra sp. Platyhelminthes Turbellaria Turbellaria Nematoda Nematoda Bryozoa Bryozoa Mollusca Gastropoda Gastropoda Mollusca Gastropoda Mesogastropoda Mesogastropoda Mollusca Gastropoda Mesogastropoda Viviparidae Campeloma Campeloma sp. Mollusca Gastropoda Mesogastropoda Viviparidae Viviparus Viviparus sp. Mollusca Gastropoda Mesogastropoda Hydrobiidae Hydrobiidae Mollusca Gastropoda Basommatophora Ancylidae Ancylidae Mollusca Gastropoda Basommatophora Ancylidae Ferrissia Ferrissia sp. -

Land Use Management System for the Nelson Mandela Metropolitan Municipality
TOWARDS A NEW LAND USE MANAGEMENT SYSTEM FOR THE NELSON MANDELA METROPOLITAN MUNICIPALITY COMPONENT 1 : ANALYSIS & POLICY DIRECTIVES PHASES 1, 2 & 3 OUTCOMES REPORT (1ST DRAFT : FEBRUARY 2005) Prepared by: Prepared for: Business Unit Manager: Housing and Land Nelson Mandela Metropolitan Municipality Mr J. van der Westhuysen i Executive Summary 1.0 INTRODUCTION 1.1 INTRODUCTION Drafting of a New Land Use Management System (LUMS) for the Nelson Mandela Metropolitan Municipality, which should include as a minimum requirement a single zoning scheme for the Metro‟s area of jurisdiction, has been identified as a priority project in the Metro‟s 2003/4 Integrated Development Plan (IDP). 1.2 PROBLEM STATEMENT With the amalgamation of the Rural Areas, Uitenhage, Despatch and Port Elizabeth, in 2000, the Nelson Mandela Metropolitan Municipality inherited twelve (12) different sets of zoning schemes. These zoning schemes are currently administered and implemented by the Municipality and in some cases delegation vests with the Department of Housing, Local Government and Traditional Affairs. These zoning schemes, some dating back to 1961, were prepared and promulgated in terms of various sets of legislation, i.e. the Land Use Planning Ordinance and the regulations promulgated in terms of the Black Communities Development Act. In many respects, these existing zoning schemes are inappropriate and outdated and therefore do not respond to current and identified future land development and conservation needs. The White Paper on Spatial Planning and Land Use Management clearly summarises shortfalls relating to land use planning and management in general: . Disparate Land Use Management Systems in Former “Race Zones” . Old and outdated Land Use Management . -

Gustafson and Sites: D. Shorti a New Species of Whirligig
Gustafson and Sites: D. shorti a new species Grey Gustafson of whirligig beetle from the U.S. Department of Biology 167 Castetter Hall Annals of the Entomological Society of MSC03 2020 America 1 University of New Mexico Albuquerque, NM 87131-0001 Phone: 303-929-9606 Email: [email protected] ********************** PREPRINT FOR OPEN ACCESS *********************** A North American biodiversity hotspot gets richer: A new species of whirligig beetle from the Southeastern Coastal Plain of the United States Grey T. Gustafson1 and Robert W. Sites2 1Museum of Southwestern Biology, Department of Biology, University of New Mexico, Albuquerque, New Mexico 87131 U.S.A 2Enns Entomology Museum, Division of Plant Sciences, University of Missouri, Columbia, Missouri 65211, U.S.A. Abstract A new species of Dineutus Macleay, 1825 is described from the Southeastern Coastal Plain of the United Sates. Habitus and aedeagus images as well as illustrations of elytral apices, protarsus, palps, and male mesopretarsal claws are provided for Dineutus shorti n. sp. and compared to those of D. discolor Aubé, 1838. The importance of the Southeastern Coastal Plain as a biodiversity hotspot and the potential conservation concern of D. shorti n. sp. also are discussed. Keywords Aquatic beetle, new species, biodiversity, conservation, Southeastern Coastal Plain Introduction Whirligig beetles are commonly encountered aquatic beetles and are well known for their whirling swimming pattern (Tucker 1969) and propensity to form aggregates of hundreds to thousands of individuals (Heinrich and Vogt 1980). These beetles are especially abundant in the southeastern United States where all four genera known to occur in the U.S. can be found (Epler 2010). -

(Tropocorixa) Jensenhaarupi (Hemiptera: Heteroptera: Corixidae: Corixini), with Ecological Notes Revista Mexicana De Biodiversidad, Vol
Revista Mexicana de Biodiversidad ISSN: 1870-3453 [email protected] Universidad Nacional Autónoma de México México Melo, María Cecilia; Scheibler, Erica Elizabeth Description of the immature stages ofSigara (Tropocorixa) jensenhaarupi (Hemiptera: Heteroptera: Corixidae: Corixini), with ecological notes Revista Mexicana de Biodiversidad, vol. 82, núm. 1, marzo, 2011, pp. 117-130 Universidad Nacional Autónoma de México Distrito Federal, México Available in: http://www.redalyc.org/articulo.oa?id=42520745011 How to cite Complete issue Scientific Information System More information about this article Network of Scientific Journals from Latin America, the Caribbean, Spain and Portugal Journal's homepage in redalyc.org Non-profit academic project, developed under the open access initiative Revista Mexicana de Biodiversidad 82: 117-130, 2011 Description of the immature stages of Sigara (Tropocorixa) jensenhaarupi (Hemiptera: Heteroptera: Corixidae: Corixini), with ecological notes Descripción de los estadios larvales de Sigara (Tropocorixa) jensenhaarupi (Heteroptera: Corixidae), con notas acerca de su ecología María Cecilia Melo1* and Erica Elizabeth Scheibler2 1Departamento Sistemática, Instituto de Limnología “R.A. Ringuelet” (ILPLA) (CCT La Plata CONICET- UNLP), C.C. 712, 1900 La Plata, Argentina. 2Laboratorio de Entomología, Instituto Argentino de Investigaciones de las Zonas Áridas (IADIZA, CCT CONICET, Mendoza), C.C. 507, 5500 Men- doza, Argentina. *Correspondent: [email protected] Abstract. Sigara (Tropocorixa) jensenhaarupi Jaczewski is the smallest species of the subgenus ranging from 4.2-4.7 mm, and it is characterized by the absence of a strigil, the small and narrow genital capsule with a short hypandrium in males, and the shape of the abdominal tergite VII in females. This species is endemic to the Patagonian subregion (Andean region) in Argentina.