Appendix J FWS and NOAA Fisheries Biological Opinions
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

2020 Coastal Massachusetts COASTSWEEP Results (People
COASTSWEEP 2020 - Cleanup Results Town Location Group Name People Pounds Miles TOTALS 703 9016.2 151.64 Arlington Mystic River near River Street 1 2 Arlington Mystic River 1 2.12 1.20 Barnstable Sandy Neck Beach Take Care Cape Cod 4 27.5 3.95 Barnstable Jublilation Way, Osterville 1 0.03 Barnstable Sandy Neck Beach Take Care Cape Cod 2 10.13 0.53 Barnstable Sandy Neck Beach Take Care Cape Cod 1 8 Barnstable Sandy Neck Beach Take Care Cape Cod 2 8.25 1.07 Barnstable Sandy Neck Beach Take Care Cape Cod 3 14.25 1.16 Barnstable Oregon Beach, Cotuit 6 30 Barnstable KalMus Park Beach 2 23.63 0.05 Barnstable Dowes Beach, East Bay Cape Cod Anti-Litter Coalition 4 25.03 0.29 Barnstable Osterville Point, Osterville Cape Cod Anti-Litter Coalition 1 3.78 0.09 Barnstable Louisburg Square, Centerville 2 Barnstable Hathaway's Ponds 2 4.1 0.52 Barnstable Hathaway's Ponds 2 5.37 0.52 Barnstable Eagle Pond, Cotuit Lily & Grace Walker 2 23.75 3.26 Beverly Corning Street SaleM Sound Coastwatch 2 0.02 Beverly Corning Street SaleM Sound Coastwatch 1 0.07 0.02 Beverly Corning Street SaleM Sound Coastwatch 1 0.03 0.02 Beverly Corning Street SaleM Sound Coastwatch 1 0.11 0.02 Beverly Corning Street SaleM Sound Coastwatch 1 0.18 0.01 Beverly Dane Street Beach SaleM Sound Coastwatch 1 0.36 0.04 Beverly Clifford Ave 2 11.46 0.03 Beverly Near David Lynch Park 1 0.43 0.03 Beverly Rice's Beach SaleM Sound Coastwatch 3 28.61 0.03 Beverly Rice's Beach SaleM Sound Coastwatch 3 1.61 Beverly Rice's Beach SaleM Sound Coastwatch 1 0.07 COASTSWEEP 2020 - Cleanup Results Town -

Rapid Formation and Degradation of Barrier Spits in Areas with Low Rates of Littoral Drift*
Marine Geology, 49 (1982) 257-278 257 Elsevier Scientific Publishing Company, Amsterdam- Printed in The Netherlands RAPID FORMATION AND DEGRADATION OF BARRIER SPITS IN AREAS WITH LOW RATES OF LITTORAL DRIFT* D.G. AUBREY and A.G. GAINES, Jr. Woods Hole Oceanographic Institution, Woods Hole, MA 02543 (U.S.A.) (Received February 8, 1982; revised and accepted April 6, 1982) ABSTRACT Aubrey, D.G. and Gaines Jr., A.G., 1982. Rapid formation and degradation of barrier spits in areas with low rates of littoral drift. Mar. Geol., 49: 257-278. A small barrier beach exposed to low-energy waves and a small tidal range (0.7 m) along Nantucket Sound, Mass., has experienced a remarkable growth phase followed by rapid attrition during the past century. In a region of low longshore-transport rates, the barrier spit elongated approximately 1.5 km from 1844 to 1954, developing beyond the baymouth, parallel to the adjacent Nantucket Sound coast. Degradation of the barrier spit was initiated by a succession of hurricanes in 1954 (Carol, Edna and Hazel). A breach opened and stabilized near the bay end of the one kilometer long inlet channel, providing direct access for exchange of baywater with Nantucket Sound, and separating the barrier beach into two nearly equal limbs. The disconnected northeast limb migrated shorewards, beginning near the 1954 inlet and progressing northeastward, filling the relict inlet channel behind it. At present, about ten percent of the northeast limb is subaerial: the rest of the limb has completely filled the former channel and disappeared. The southwest limb of the barrier beach has migrated shoreward, but otherwise has not changed significantly since the breach. -

Ipswich Where to Go • What to See • What to Do
FINAL-1 Wed, Jun 21, 2017 8:03:55 PM DESTINATION IPSWICH WHERE TO GO • WHAT TO SEE • WHAT TO DO Nicole Goodhue Boyd Nicole The Salem News PHOTO/ FINAL-1 Wed, Jun 21, 2017 8:03:57 PM S2 • Friday, June 23, 2017 June • Friday, DESTINATION IPSWICH DESTINATION Trust in Our Family Business The Salem News • News Salem The Marcorelle’s Fine Wine, Liquor & Beer Specializing in beverage catering, functions and delivery since 1935. 30 Central Street, Ipswich, MA 01938, Phone: 978-356-5400 Proud retailer of Ipswich Ale Brewery products Visit ipswichalebrewery.com for brewery tour & restaurant hours. FINAL-1 Wed, Jun 21, 2017 8:03:58 PM S3 The Salem News • News Salem The Family Owned & Operated Since 1922 IPSWICH DESTINATION • Send someone flowers, make someone happy • Colorful Hanging Baskets and 23, 2017 June • Friday, colorful flowering plants for all summer beauty • Annuals and Perennials galore • Fun selection of quality succulents & air plants • Walk in cut flower cooler • Creative Floral Arrangements • One of a Kind Gifts & Cards Friend us on www.gordonblooms.com 24 Essex Rd. l Ipswich, MA l 978.356.2955 FINAL-1 Wed, Jun 21, 2017 8:03:58 PM S4 RECREATION • Friday, June 23, 2017 June • Friday, DESTINATION IPSWICH DESTINATION The Salem News • News Salem The File photos The rooftop views from the Great House at the Crane Estate Crane Beach is one of the most popular go-to spots for playing on the sand and in the water. include the “allee” that leads to the Atlantic Ocean. Explore the sprawling waterways and trails Visitors looking to get through the end of October. -

Cape Poge Wildlife Refuge, Leland Beach, Wasque Point, and Norton Point Beach Edgartown
Impact Avoidance and Minimization Plan: Cape Poge Wildlife Refuge, Leland Beach, Wasque Point, and Norton Point Beach Edgartown, Martha’s Vineyard January 2020 The Trustees of Reservations 200 High Street Boston, MA 02110 Table of Contents 1. Site Description 1.a Maps……………………………………………………………………………………………………………………… 1 1.b Description of site…………………………………………………………………………………………………. 3 1.c habitat and management………………………………………………………………………………………. 5 1.d Plover breeding a productivity………………………………………………………..…………………….. 6 2. Responsible Staff 2.a Staff biographies……………………………………………………………………………………………………. 8 3. Beach Management 3.a.i Recreational Activities………………………………………………………………………………………… 9 3.a.ii Parking and Roads……………………………………………………………………………………….……. 9 3.a.iii Beach cleaning and refuse management…………………………………..……………………. 10 3.a.iv Rules and regulations…………………………………………………………………………….……….... 10 3.a.v Law enforcement…………………………………………………………………………….………………… 10 3.a.vi Other management……………………………………………………………………………………………. 10 3.a.vi Piping plover management……………………………………………………………………………….. 10 4. Covered Activities 4.1.a OSV use in vicinity of piping plover chicks…………………………………………………………….. 12 4.1.b Reduced symbolic fencing……………………………………………………………………………………. 15 4.1.c Reduced proactive symbolic fencing……………………………………………………………………… 16 4.2 Contingency Plan…………………………………………………………………………………….……………. 18 4.3 Violations………………………………………………………………………………………………………………… 18 4.4 Self-escort program reporting………………………………………………………………………………… 18 5. Budget…………………………………………………………………………………………………………………………. -

City of Gloucester Open Space and Recreation Plan 2010 - 2017
City of Gloucester Open Space and Recreation Plan 2010 - 2017 Updated from plan dated 1998 - 2003 The City of Gloucester, Carolyn Kirk, Mayor Open Space & Recreation Committee Daniel Morris (Chair) Noel Mann Charles Crowley Susan Hedman John McElhenny Dean Murray Kathy Leahy (thru 5/2010) Patti Amaral (post 6/2010) Gloucester Open Space & Recreation Plan 2010-2017 3-31-2011 Final OPEN SPACE AND RECREATION PLAN Table of Contents Letter from the Open Space and Recreation Com m ittee Section 1: PLAN SUM M ARY ................................................................. 1 Section 2: INTRODUCTION … … … … … … … … … … … … … … … … … … … … … ..5 A. Statem ent of Purpose: W hy W rite This Plan? B. Planning Process and Public Participation C. Environm ental Justice Com m unity Outreach D. How This Plan Qualifies Gloucester for Grants Section 3: COM M UNITY SETTING … … … … … … … … … … … … … … … … … .. 10 A. Regional Context: Gloucester and the Cape Ann Peninsula B. History of Gloucester C. Population Characteristics of Gloucester D. Growth and Developm ent Pattern E. Regional Open Space & Related Plans Section 4: ENVIRONM ENTAL INVENTORY AND ANALYSIS … … … … … … 31 A. Geology, Soils, and Topography B. Landscape Character C. W ater Resources D. Vegetation E. Fisheries and W ildlife F. Scenic Resources and Unique Environm ents G. Environm ental Challenges Section 5: INVENTORY OF LANDS OF CONSERVATION AND … … … … … .60 RECREATION INTEREST A. Private Parcels B. Public and Nonprofit Parcels 1. Active Recreation Areas: Beaches, Fields, Parks & Playgrounds 2. Open Land: Conservation, W atershed and W oodland Areas 3. Public Landings Section 6: GLOUCESTER’S VISION FOR OPEN SPACE & RECREATION ..79 A. Description of Process B. Statem ent of Open Space and Recreation Goals Section 7: W HAT GLOUCESTER NEEDS … … … … … … … … … … … … … … … . -
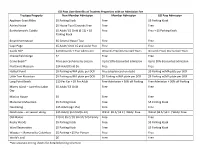
GO Pass User Benefits at Trustees Properties with an Admission Fee
GO Pass User Benefits at Trustees Properties with an Admission Fee Trustees Property Non-Member Admission Member Admission GO Pass Admission Appleton Grass Rides $5 Parking Kiosk Free $5 Parking Kiosk Ashley House $5 House Tour/Grounds Free Free Free Bartholomew’s Cobble $5 Adult/ $1 Child (6-12) + $5 Free Free + $5 Parking Kiosk Parking Kiosk Bryant Homestead $5 General House Tour Free Free Cape Poge $5 Adult/ Child 15 and under free Free Free Castle Hill* $10 Grounds + Tour Admission Grounds Free/Discounted Tours Grounds Free/ Discounted Tours Chesterfield Gorge $2 Free Free Crane Beach* Price per car/varies by season Up to 50% discounted admission Up to 50% discounted admission Fruitlands Museum $14 Adult/Child $6 Free Free Halibut Point $5 Parking w/MA plate per DCR Free (display card on dash) $5 Parking w/MA plate per DCR Little Tom Mountain $5 Parking w/MA plate per DCR $5 Parking w/MA plate per DCR $5 Parking w/MA plate per DCR Long Point Beach $10 Per Car + $5 Per Adult Free Admission + 50% off Parking Free Admission + 50% off Parking Misery Island – June thru Labor $5 Adult/ $3 Child Free Free Day Mission House $5 Free Free Monument Mountain $5 Parking Kiosk Free $5 Parking Kiosk Naumkeag $15 Adult (age 15+) Free Free Notchview – on season skiing $15 Adult/ $6 Child (6-12) Wknd: $8 A/ $3 C | Wkdy: Free Wknd: $8 A/ $3 C | Wkdy: Free Old Manse $10 A/ $5 C/ $9 SR+ST/ $25 Family Free Free Rocky Woods $5 Parking Kiosk Free $5 Parking Kiosk Ward Reservation $5 Parking Kiosk Free $5 Parking Kiosk Wasque – Memorial to Columbus $5 Parking + $5 Per Person Free Free World’s End $6 Free Free *See separate pricing sheets for detailed pricing structure . -

Special Places : a Newsletter of the Trustees of Reservations
The Trustees of Reservations Conserving the Massachusetts Landscape Since 1891 SPECIAL PLACES Volume 7, No. 4 f Fall 1999 A QUARTERLY NEWSLETTER For Members and Donors of The Trustees of Reservations WHAT'S GOING ON HERE? See page 7 (center right) for the ecology scoop! Ecology: A Natural Part of Our Stewardship of Special Places It's 10:00 P.M. at Bartholomew's Cobble and ecologist Don in Massachusetts—from coastal heathlands to barrier Reid is standing in the pouring rain with a field guide in beaches, from rocky summits to boreal forests. hand. "What's that sound?" he mutters to himself. Don tilts The complex array of natural communities, and the his head as he strains to wildlife they support, hear above the splatter of comprise the ecological raindrops the subtle snoring resources of which we are of a frog from the nearby stewards. By scientifically wetlands. He wonders... is it evaluating these resources the Northern Pickerel Frog and applying sound or the Southern Pickerel practices to their manage- Frog? "Which one is it?" he ment, we help to conserve asks. "Why does it matter?" and, where possible, enhance you may ask. (Find out the their values, not just for answer on page 3!) the benefit of the reservation Don is one of a small itself but for the Massa- group of professional chusetts landscape in all ecologists working its diversity. throughout the state to While a major part make sure that the natural of our management, The systems associated with our Trustees' work in ecology TheTrustees EcologyTeam (left to right) Lisa Vernegaard, Vin Antil, Wayne 83 reservations remain Castonguay, Sally Matkovich, Don Reid, Russ Hopping, and Lloyd Raleigh. -

TOWN of MASHPEE BLUE PAGES a Citizens’ Guide to Protecting Cape Cod Waters
TOWN OF MASHPEE BLUE PAGES A Citizens’ Guide to Protecting Cape Cod Waters Shannon Cushing, Grade 11 Acknowledgements Acknowledgements This information is reprinted from the Island Blue Pages, courtesy of the Martha’s Vineyard Shellfish Group and the WampanoagThis information Tribe ofis Aquinnah.reprinted from For the a complete Island Blue version Pages of, courtesy the Island of theBlue Martha’s Pages, visit Vineyard the website Shellfish www.islandbluepages.org Group and the . or contactWampanoag tbe Martha’s Tribe Vineyardof Aquinnah. Shellfish For a Groupcomplete at 508version-693-0391. of the TheIsland Island Blue BluePages Pages, visit isthe an website adaptation, www.islandbluepages.org with permission, of the. or Pugetcontact Soundbook tbe Martha’s, a game Vineyard plan for Shellfish maintaining Group the at health508-693-0391. of our sister The Island estuary Blue on Pagesthe West is an Coast. adaptation, To learn with more permission, about the of the Puget Soundbook, a game plan for maintaining the health of our sister estuary on the West Coast. To learn more about the original project and the inspiration for the Blue Pages, visit www.forsea.org/pugetsoundbook/ original project and the inspiration for the Blue Pages, visit www.forsea.org/pugetsoundbook/ Thanks to Jim Kolb and Diane Bressler, the creators of the Puget Soundbook, which continues to inspire us with its words and Thanks to Jim Kolb and Diane Bressler, the creators of the Puget Soundbook, which continues to inspire us with its words and illustrations. illustrations. The Town of Mashpee, with permission, undertook the task of adapting the Orleans Blue Pages to reflect conditions specific to The Town of Mashpee, with permission, undertook the task of adapting the Orleans Blue Pages to reflect conditions specific to Mashpee. -

Adaptation Strategies for the Great Marsh Region
Ranger Poole/USFWS CHAPTER 4 Adaptation Strategies for the Great Marsh Region Assessing vulnerability is the first step in generating adaptation options to increase resilience and reduce vulnerability. Understanding why an asset is vulnerable is especially critical to thinking about adaptation and in particular, identifying adaptation options that can address one or more of the three components of vulnerability (i.e. exposure, sensitivity, and adaptive capacity). Furthermore, while vulnerability assessments provide the context necessary for identifying important issues to consider when designing adaptation strategies, the identification of “key vulnerabilities” can help steer the generation of adaptation options in a direction that focuses on the most critical issues. 376 This chapter outlines a range of adaptation strategies identified through the Community Adaptation Planning Process (see Chapter 2). The following strategies and recommendations are broken into two categories: regional strategies and town-specific strategies. Successful short and long-term implementation of all of these recommendations will require an extensive amount of intra- and inter- municipal cooperation, regional collaboration, and ongoing environmental research and monitoring. The Great Marsh Region is fortunate to have a wide diversity of organizations, agencies, and municipalities working to protect and restore the Great Marsh. However, these efforts will need to be continually strengthened to achieve the degree of change and level of project implementation recommended in this report. 376 Stein, B.A., P. Glick, N. Edelson, and A. Staudt (eds.), Climate-Smart Conservation: Putting Adaptation Principles into Practice (Reston, VA: National Wildlife Federation, 2014), 120 Ch 4. Adaptation Strategies for the Great Marsh Region | 124 4.1 Regional Strategies and Recommendations This section highlights adaptation strategies that should be adopted to reduce vulnerability on a regional scale. -

MDPH Beaches Annual Report 2008
Marine and Freshwater Beach Testing in Massachusetts Annual Report: 2008 Season Massachusetts Department of Public Health Bureau of Environmental Health Environmental Toxicology Program http://www.mass.gov/dph/topics/beaches.htm July 2009 PART ONE: THE MDPH/BEH BEACHES PROJECT 3 I. Overview ......................................................................................................5 II. Background ..................................................................................................6 A. Beach Water Quality & Health: the need for testing......................................................... 6 B. Establishment of the MDPH/BEHP Beaches Project ....................................................... 6 III. Beach Water Quality Monitoring...................................................................8 A. Sample collection..............................................................................................................8 B. Sample analysis................................................................................................................9 1. The MDPH contract laboratory program ...................................................................... 9 2. The use of indicators .................................................................................................... 9 3. Enterococci................................................................................................................... 10 4. E. coli........................................................................................................................... -

2019 Coastal Massachusetts COASTSWEEP Results
Results of the COASTSWEEP 2019 Cleanups Organization Coordinator Site(s) Town(s) People Pounds Miles Sustainable Practices Karla Cabral Craigville Beach, Covells Beach Barnstable 29 46 0.50 Sustainable Practices Karla Cabral Kalmus Beach Barnstable 9 18 0.50 Town of Barnstable Susan Brinckerhoff Millway Beach Barnstable 23 2 0.50 Town of Barnstable Betsy Wheeler Rendezvous Lane Barnstable 1 10 0.50 Town of Barnstable Robert Parsons Scudders Lane Barnstable 13 50 1 Town of Barnstable Fred Stepanis Sandy Neck Beach Barnstable 37 100 6.00 Sustainable Practices Patrick Otton Town Beach Barnstable 13 40 0.75 Rotary Club of Osterville Anke Rudy Dowes Beach Barnstable 38 50 2.00 Wellfleet Recycling Committee and Sustainable Christine Shreves Duck Harbor Barnstable 11 25 0.25 Practices Salem Sound Coastwatch Margaret Duffy Dane Street Beach Beverly 24 30 0.50 Endicott College Lori Mitchener Endicott College Beach Beverly 2 7 0.25 Salem Sound Coastwatch Margaret Duffy West Beach Beverly 8 13 1.00 Glen Urquhart School Laura Doyle Dane Street Beach Beverly 25 10 0.25 Salem Sound Coastwatch Margaret Duffy McPherson Park Beverly 8 20 0.25 Results of the COASTSWEEP 2019 Cleanups Organization Coordinator Site(s) Town(s) People Pounds Miles Salem Sound Coastwatch Margaret Duffy Sandy Point Beverly 7 60 0.50 Urban Harbors Institute and the School for the Kimberly Starbuck UMass Boston Beach Boston 100 200 2.00 Environment at UMass Boston Tenacre Country Day School Leah Staffier Carson Beach Boston 30 30 0.50 ClearView Healthcare Partners Mara Blumenstein -

E. Fisheries and Wildlife
E. Fisheries and Wildlife Until recent decades, the vast majority of Mashpee’s territory was the domain not of man, but of beast. Our woods were only occasionally broken by a roadway, or a few homes, or a farmer’s fields. The hunting was good. Our clear lakes were famous for their fishing. Our streams flowed clean to pristine coastal bays teeming with fish and shellfish that helped feed and support many families. Much has changed with the explosive development of the last fifty years, and much of our wildlife has disappeared along with the natural habitat that supported it. However, much remains for our enjoyment and safekeeping. In this section we will look at Mashpee’s fish and shellfish, its mammals, birds and insects, important wildlife movement corridors and those species living in our town which are among the last of their kind. 1. Finfish Mashpee hosts four types of fin fisheries: fresh water ponds, rivers and streams, estuaries and coastal ponds and the open ocean. Our four large ponds provide some of the best fishing in the state. 203-acre Ashumet Pond, 317- acre Johns Pond and 729-acre Mashpee-Wakeby Pond are all cold water fisheries stocked with brown, brook and rainbow trout. In the last century, such famous anglers as Daniel Webster, President Grover Cleveland and the famous actor Joseph Jefferson looked forward to their fishing expeditions to Mashpee, while local residents looked forward to the income provided serving as guides to those and other wealthy gentlemen. Ashumet and Johns Ponds are also noted for their smallmouth bass, while Mashpee-Wakeby provides not only the smallmouth, but also chain pickerel, white perch and yellow perch.