Scientific Advice on Matters Related to the Management of Seal Populations: 2008
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

FSG Report 2010
Forth Seabird Group Forth Islands Bird Report 2010 Compiled by Bill Bruce April 2011 SEABIRD SPECIES SUMMARIES 2010 Note: AOS = Apparently occupied sites; AON = Apparently occupied nests; AOB = Apparently occupied burrows; AOT = Apparently occupied territories FULMAR GREAT BLACK-BACKED GULL Overall, total numbers are very similar to last year On Fidra, Carr Craig and Inchmickery numbers (up 18 birds, 1%) are the same as last year while all other islands show a decrease. Overall numbers are down by 12 CORMORANT AON (17%) On Craigleith and Inchkeith breeding numbers LESSER BLACK-BACKED GULL & are exactly the same as last year. For the second HERRING GULL year none were breeding on Haystack while on Carr Craig and Lamb numbers are down. Overall These gulls were counted on Inchkeith, the first numbers are down by 21 AON or 8%. count since 2002. This showed that were 2620 AON / 3720 AOT for herring gulls and 2670 AON SHAG / 3500 AOT for LBB gulls Biggest increases are on Lamb (+39 AON, 52%) KITTIWAKE and Fidra (+45 AON, 28%) while the biggest decrease is on Inchkeith (-32 AON, -20%). The With the exception of 2009, numbers of AON for total for all islands is up 135 AON (12%), which all islands have fluctuated by approx. ±4% for a continues the general increase since numbers number of years. The May Isle in 2009 was lower halved between 2004 and 2005. (21% down compared to 2008) than this general trend would expect but numbers have recovered GANNET this year and are slightly up on the 2007 count. Not counted this year. -

List of Lights Radio Aids and Fog Signals 2011
PUB. 114 LIST OF LIGHTS RADIO AIDS AND FOG SIGNALS 2011 BRITISH ISLES, ENGLISH CHANNEL AND NORTH SEA IMPORTANT THIS PUBLICATION SHOULD BE CORRECTED EACH WEEK FROM THE NOTICE TO MARINERS Prepared and published by the NATIONAL GEOSPATIAL-INTELLIGENCE AGENCY Bethesda, MD © COPYRIGHT 2011 BY THE UNITED STATES GOVERNMENT. NO COPYRIGHT CLAIMED UNDER TITLE 17 U.S.C. *7642014007536* NSN 7642014007536 NGA REF. NO. LLPUB114 LIST OF LIGHTS LIMITS NATIONAL GEOSPATIAL-INTELLIGENCE AGENCY PREFACE The 2011 edition of Pub. 114, List of Lights, Radio Aids and Fog Signals for the British Isles, English Channel and North Sea, cancels the previous edition of Pub. 114. This edition contains information available to the National Geospatial-Intelligence Agency (NGA) up to 2 April 2011, including Notice to Mariners No. 14 of 2011. A summary of corrections subsequent to the above date will be in Section II of the Notice to Mariners which announced the issuance of this publication. In the interval between new editions, corrective information affecting this publication will be published in the Notice to Mariners and must be applied in order to keep this publication current. Nothing in the manner of presentation of information in this publication or in the arrangement of material implies endorsement or acceptance by NGA in matters affecting the status and boundaries of States and Territories. RECORD OF CORRECTIONS PUBLISHED IN WEEKLY NOTICE TO MARINERS NOTICE TO MARINERS YEAR 2011 YEAR 2012 1........ 14........ 27........ 40........ 1........ 14........ 27........ 40........ 2........ 15........ 28........ 41........ 2........ 15........ 28........ 41........ 3........ 16........ 29........ 42........ 3........ 16........ 29........ 42........ 4....... -

The Case for a Marine Act for Scotland the Tangle of the Forth
The Case for a Marine Act for Scotland The Tangle of the Forth © WWF Scotland For more information contact: WWF Scotland Little Dunkeld Dunkeld Perthshire PH8 0AD t: 01350 728200 f: 01350 728201 The Case for a Marine Act for Scotland wwf.org.uk/scotland COTLAND’S incredibly Scotland’s territorial rich marine environment is waters cover 53 per cent of Designed by Ian Kirkwood Design S one of the most diverse in its total terrestrial and marine www.ik-design.co.uk Europe supporting an array of wildlife surface area Printed by Woods of Perth and habitats, many of international on recycled paper importance, some unique to Scottish Scotland’s marine and WWF-UK registered charity number 1081274 waters. Playing host to over twenty estuarine environment A company limited by guarantee species of whales and dolphins, contributes £4 billion to number 4016274 the world’s second largest fish - the Scotland’s £64 billion GDP Panda symbol © 1986 WWF – basking shark, the largest gannet World Wide Fund for Nature colony in the world and internationally 5.5 million passengers and (formerly World Wildlife Fund) ® WWF registered trademark important numbers of seabirds and seals 90 million tonnes of freight Scotland’s seas also contain amazing pass through Scottish ports deepwater coral reefs, anemones and starfish. The rugged coastline is 70 per cent of Scotland’s characterised by uniquely varied habitats population of 5 million live including steep shelving sea cliffs, sandy within 0km of the coast and beaches and majestic sea lochs. All of 20 per cent within km these combined represent one of Scotland’s greatest 25 per cent of Scottish Scotland has over economic and aesthetic business, accounting for 11,000km of coastline, assets. -

Layout 1 Copy
STACK ROCK 2020 An illustrated guide to sea stack climbing in the UK & Ireland - Old Harry - - Old Man of Stoer - - Am Buachaille - - The Maiden - - The Old Man of Hoy - - over 200 more - Edition I - version 1 - 13th March 1994. Web Edition - version 1 - December 1996. Web Edition - version 2 - January 1998. Edition 2 - version 3 - January 2002. Edition 3 - version 1 - May 2019. Edition 4 - version 1 - January 2020. Compiler Chris Mellor, 4 Barnfield Avenue, Shirley, Croydon, Surrey, CR0 8SE. Tel: 0208 662 1176 – E-mail: [email protected]. Send in amendments, corrections and queries by e-mail. ISBN - 1-899098-05-4 Acknowledgements Denis Crampton for enduring several discussions in which the concept of this book was developed. Also Duncan Hornby for information on Dorset’s Old Harry stacks and Mick Fowler for much help with some of his southern and northern stack attacks. Mike Vetterlein contributed indirectly as have Rick Cummins of Rock Addiction, Rab Anderson and Bruce Kerr. Andy Long from Lerwick, Shetland. has contributed directly with a lot of the hard information about Shetland. Thanks are also due to Margaret of the Alpine Club library for assistance in looking up old journals. In late 1996 Ben Linton, Ed Lynch-Bell and Ian Brodrick undertook the mammoth scanning and OCR exercise needed to transfer the paper text back into computer form after the original electronic version was lost in a disk crash. This was done in order to create a world-wide web version of the guide. Mike Caine of the Manx Fell and Rock Club then helped with route information from his Manx climbing web site. -

Records of Species and Subspecies Recorded in Scotland on up to 20 Occasions
Records of species and subspecies recorded in Scotland on up to 20 occasions In 1993 SOC Council delegated to The Scottish Birds Records Committee (SBRC) responsibility for maintaining the Scottish List (list of all species and subspecies of wild birds recorded in Scotland). In turn, SBRC appointed a subcommittee to carry out this function. Current members are Dave Clugston, Ron Forrester, Angus Hogg, Bob McGowan Chris McInerny and Roger Riddington. In 1996, Peter Gordon and David Clugston, on behalf of SBRC, produced a list of records of species recorded in Scotland on up to 5 occasions (Gordon & Clugston 1996). Subsequently, SBRC decided to expand this list to include all acceptable records of species recorded on up to 20 occasions, and to incorporate subspecies with a similar number of records (Andrews & Naylor 2002). The last occasion that a complete list of records appeared in print was in The Birds of Scotland, which included all records up until 2004 (Forrester et al. 2007). During the period from 2002 until 2013, amendments and updates to the list of records appeared regularly as part of SBRC’s Scottish List Subcommittee’s reports in Scottish Birds. Since 2014 these records have appear on the SOC’s website, a significant advantage being that the entire list of all records for such species can be viewed together (Forrester 2014). The Scottish List Subcommittee are now updating the list annually. The current update includes records from the British Birds Rarities Committee’s Report on rare birds in Great Britain in 2015 (Hudson 2016) and SBRC’s Report on rare birds in Scotland, 2015 (McGowan & McInerny 2017). -

Talking About Heritage
Talking about heritage Draft guidance for consultation September 2020 1 Introduction Heritage is everywhere and it means different things to different people. This guide is all about exploring and talking about heritage, so we’ve included some of the things that people have said to us when we’ve asked them, ‘What’s your heritage?’ Heritage to me is everything in Scotland’s history. It’s not just buildings but everything that’s passed down like songs, stories, myths. Perthshire ‘What’s Your Heritage’ workshop. Your heritage might be the physical places that you know and love – your favourite music venue, your local park, a ruined castle you’ve explored, or the landscapes you picture when you think of home. Your heritage could also be your working life, the stories you were told as a child, the language you speak with your family, the music or traditions you remember from an important time in your life. Heritage can inspire different emotions, both positive and negative. It can be special to people for lots of different reasons. Here are a few: • It’s beautiful. • It’s what I think of when I picture home. • It’s part of who I am • I can feel the spirits, my history. • It’s where I walk my dog. • It’s an amazing insight into my past. • It’s my home town and it reminds me of my family. Heritage can help to us to feel connected. It might be to a community, a place, or a shared past. It reflects different viewpoints across cultures and generations and is key to local distinctiveness and identity. -

Greenland Barnacle 2003 Census Final
GREENLAND BARNACLE GEESE BRANTA LEUCOPSIS IN BRITAIN AND IRELAND: RESULTS OF THE INTERNATIONAL CENSUS, MARCH 2003 WWT Report Authors Jenny Worden, Carl Mitchell, Oscar Merne & Peter Cranswick March 2004 Published by: The Wildfowl & Wetlands Trust Slimbridge Gloucestershire GL2 7BT T 01453 891900 F 01453 891901 E [email protected] Reg. charity no. 1030884 © The Wildfowl & Wetlands Trust All rights reserved. No part of this document may be reproduced, stored in a retrieval system or transmitted, in any form or by any means, electronic, mechanical, photocopying, recording or otherwise without the prior permission of WWT. This publication should be cited as: Worden, J, CR Mitchell, OJ Merne & PA Cranswick. 2004. Greenland Barnacle Geese Branta leucopsis in Britain and Ireland: results of the international census, March 2003 . The Wildfowl & Wetlands Trust, Slimbridge. gg CONTENTS Summary v 1 Introduction 6 2 Methods 7 3 Results 8 4 Discussion 13 4.1 Census total and accuracy 13 4.2 Long-term trend and distribution 13 4.3 Internationally and nationally important sites 17 4.4 Future recommendations 19 5 Acknowledgements 20 6 References 21 Appendices 22 ggg SUMMARY Between 1959 and 2003, eleven full international surveys of the Greenland population of Barnacle Geese have been conducted at wintering sites in Ireland and Scotland using a combination of aerial survey and ground counts. This report presents the results of the 2003 census, conducted between 27th and 31 March 2003 surveying a total of 323 islands and mainland sites along the west and north coasts of Scotland and Ireland. In Ireland, 30 sites were found to hold 9,034 Greenland Barnacle Geese and in Scotland, 35 sites were found to hold 47,256. -

The Misty Isle of Skye : Its Scenery, Its People, Its Story
THE LIBRARY OF THE UNIVERSITY OF CALIFORNIA LOS ANGELES c.'^.cjy- U^';' D Cfi < 2 H O THE MISTY ISLE OF SKYE ITS SCENERY, ITS PEOPLE, ITS STORY BY J. A. MACCULLOCH EDINBURGH AND LONDON OLIPHANT ANDERSON & FERRIER 1905 Jerusalem, Athens, and Rome, I would see them before I die ! But I'd rather not see any one of the three, 'Plan be exiled for ever from Skye ! " Lovest thou mountains great, Peaks to the clouds that soar, Corrie and fell where eagles dwell, And cataracts dash evermore? Lovest thou green grassy glades. By the sunshine sweetly kist, Murmuring waves, and echoing caves? Then go to the Isle of Mist." Sheriff Nicolson. DA 15 To MACLEOD OF MACLEOD, C.M.G. Dear MacLeod, It is fitting that I should dedicate this book to you. You have been interested in its making and in its publica- tion, and how fiattering that is to an author s vanity / And what chief is there who is so beloved of his clansmen all over the world as you, or whose fiame is such a household word in dear old Skye as is yours ? A book about Skye should recognise these things, and so I inscribe your name on this page. Your Sincere Friend, THE A UTHOR. 8G54S7 EXILED FROM SKYE. The sun shines on the ocean, And the heavens are bhie and high, But the clouds hang- grey and lowering O'er the misty Isle of Skye. I hear the blue-bird singing, And the starling's mellow cry, But t4eve the peewit's screaming In the distant Isle of Skye. -

Download Download
II.—An Account of St Columbd's Abbey, Inchcolm. Accompanied with Plans, ^c.1 (Plates IV.-VL) By THOMAS ARNOLD, Esq., Architect, M.R.LB.A, Lond. [Communicated January 11, 1869, with an Introductory Note.] NEAR the northern shores of the Firth of Forth, and within sight of Edin- burgh, lies the island anciently known as Emona, and in later times as Inchcolm, the island of St Columba. It is of very small extent, scarcely over half a mile in length, and 400 feet in width at its broadest part. The tide of commerce and busy life which ebbs and flows around has left the little inch in a solitude as profound as if it gemmed the bosom of some Highland loch, a solitude which impresses itself deeply on the stranger who comes to gaze on its ruined, deserted, and forgotten Abbey. Few even of those who visit the island from the beautiful village of Aberdour, close to it, know anything of its history, and as few out of sight of the island know of its existence at all. But although now little known beyond the shores of the Forth, Inchcolm formerly held a high place in the veneration of the Scottish people as the cradle of the religious life of the surrounding districts, and was second only to lona as a holy isle in whose sacred soil it was the desire of many generations to be buried. It numbered amongst its abbots men of high position and learning. Noble benefactors enriched it with broad lands and rich gifts, and its history and remains, like the strata of some old mountain, bear the marks of every great wave of life which has passed over our country. -

Marine Protected Areas and Large-Scale Features. Position Paper
Marine Protected Areas and Large-Scale Features. Position paper Summary This document considers the role of large-scale features in developing the Marine Protected Area (MPA) network in Scotland’s seas. It describes each of the large-scale features, outlines the approach to their inclusion in the process, and considers the possible MPAs and MPA search locations which contain these features in relation to the evidence available. Five large-scale features are included on the list of MPA search features: seamounts; continental slope; shelf deeps; shelf banks and mounds; and fronts. These features have been included to represent areas of potential wider significance to the overall health and biodiversity of Scotland’s seas in the development of the MPA network. Specific examples of large-scale features have only been included in possible MPAs and MPA search locations where evidence is available to suggest that they contribute to ecosystem function, for instance in terms of playing a key supporting role within the site or more widely; or providing functional links within the site; or in supporting linkages within the network and wider seas. It is anticipated that MPAs may be able to provide direct protection for large-scale features through management of pressures that have implications for their extent, structure and distribution within the site and thereby their wider function. Draft conservation objectives are to ‘conserve’ the function of large scale features included as protected features. In total 13 areas (10 possible MPAs and 3 MPA search locations) have been identified for large-scale features, where evidence is available to suggest they play a role in supporting ecological function. -
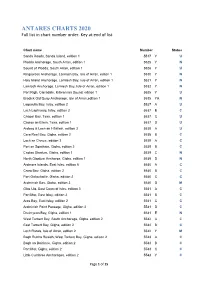
ANTARES CHARTS 2020 Full List in Chart Number Order
ANTARES CHARTS 2020 Full list in chart number order. Key at end of list Chart name Number Status Sanda Roads, Sanda Island, edition 1 5517 Y U Pladda Anchorage, South Arran, edition 1 5525 Y N Sound of Pladda, South Arran, edition 1 5526 Y U Kingscross Anchorage, Lamlash Bay, Isle of Arran, editon 1 5530 Y N Holy Island Anchorage, Lamlash Bay, Isle of Arran, edition 1 5531 Y N Lamlash Anchorage, Lamlash Bay, Isle of Arran, edition 1 5532 Y N Port Righ, Carradale, Kilbrannan Sound, edition 1 5535 Y U Brodick Old Quay Anchorage, Isle of Arran,edition 1 5535 YA N Lagavulin Bay, Islay, edition 2 5537 A U Loch Laphroaig, Islay, edition 2 5537 B C Chapel Bay, Texa, edition 1 5537 C U Caolas an Eilein, Texa, edition 1 5537 D U Ardbeg & Loch an t-Sailein, edition 3 5538 A U Cara Reef Bay, Gigha, edition 2 5538 B C Loch an Chnuic, edition 3 5539 A C Port an Sgiathain, Gigha, edition 2 5539 B C Caolas Gigalum, Gigha, edition 1 5539 C N North Gigalum Anchorge, Gigha, edition 1 5539 D N Ardmore Islands, East Islay, edition 5 5540 A C Craro Bay, Gigha, edition 2 5540 B C Port Gallochoille, Gigha, edition 2 5540 C C Ardminish Bay, Gigha, edition 3 5540 D M Glas Uig, East Coast of Islay, edition 3 5541 A C Port Mor, East Islay, edition 2 5541 B C Aros Bay, East Islay, edition 2 5541 C C Ardminish Point Passage, Gigha, edition 2 5541 D C Druimyeon Bay, Gigha, edition 1 5541 E N West Tarbert Bay, South Anchorage, Gigha, edition 2 5542 A C East Tarbert Bay, Gigha, edition 2 5542 B C Loch Ranza, Isle of Arran, edition 2 5542 Y M Bagh Rubha Ruaidh, West Tarbert -

Orkney and Shetland North Suthe
Draft Regional Locational Guidance – Wave Energy in Scottish Waters 3 NORTH The North Region three Scoping Areas of Search. The Scoping Areas of Search are: Orkney and Shetland North Sutherland Coast North West of Cape Wrath 3.1 Resource Orkney and Shetland 3.1.1 Wave resource is strong in the sea space that holds the wave area of search between Orkney and Shetland; the mean annual wave power densities that can be expected at this area of search reach a maximum of 40.5 kWatt m-1 and a minimum of 18.76 kWatt m-2. Mean annual wind power densities are of 34.4 5 kWatt m-1 in this area. 3.1.2 Summer mean wave power density is of 10.9 kWatt m-1 and winter mean power density is six times that at 59 kWatt m-1 . 3.1.3 Annual mean significant wave height is 2.48 m. In the summer the mean significant wave height is 1.63 m and the same figure for winter is 3.36 m . 3.1.4 Within the area of search the resource is higher towards the north-western part and decreases in the areas closer to the shoreline. North West of Cape Wrath & North Sutherland Coast 3.1.5 Wave resource is strong in the wave area of search at the North West of Cape Wrath, the karger of the two areas included in the map. The mean annual wave power densities that can be expected reach a maximum of 46.04 kWatt m-1 and a minimum of 27.59 kWatt m-2.