The Subterranean Asellids (Caecidoted) of Illinois (Crustacea: Isopoda: Asellidae)
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

A Conservation Focused Inventory of Subterranean Invertebrates of the Southwest Illinois Karst
Julian J. Lewis, Philip Moss, Diane Tecic, and Matthew E. Nelson - A conservation focused inventory of subterranean invertebrates of the southwest Illinois Karst. Journal of Cave and Karst Studies, v. 65, n. 1, p. 9-21. A CONSERVATION FOCUSED INVENTORY OF SUBTERRANEAN INVERTEBRATES OF THE SOUTHWESTERN ILLINOIS KARST JULIAN J. LEWIS J. Lewis and Associates, Biological Consulting, 217 W. Carter Avenue, Clarksville, IN 47129 USA PHILIP MOSS Ozark Underground Laboratory, 1572 Aley Lane, Protem, MO 65733 USA DIANE TECIC Natural Heritage Regional Administrator, 4521 Alton Commerce Parkwary, Alton, IL 62025 USA MATTHEW E. NELSON formerly The Nature Conservancy; current 7401 Placer Run, Fort Wayne, IN 46815 USA In 1998-1999 The Nature Conservancy conducted a bioinventory of caves in Monroe and St. Clair coun- ties in southwestern Illinois. This karst area comprises a small section of the Ozark Plateau isolated from the Missouri Ozarks by the Mississippi River. In the 71 sites that were sampled, 41 species thought to be globally rare were found and were assigned state (S) and global (G) ranks of rarity for conservation use. The list includes 10 species considered to be new to science and 12 species previously unreported from Illinois. Twenty four taxa were classified as obligate subterranean species, including four endemic species: the pseudoscorpion Mundochthonius cavernicolus, the amphipod Gammarus acherondytes, the milliped Chaetaspis sp. (undescribed), and the dipluran Eumesocampa sp. (undescribed). Gammarus acherondytes, recently listed as an endangered species, was found in six previously unsampled caves. All sites were rank-ordered according to the number of global and state rare species. The greatest single site diversity was found in Fogelpole Cave with 18 global and 20 state rare species. -

Downloaded from Brill.Com10/02/2021 05:23:45PM Via Free Access 24 R
Contributions to Zoology, 70 (1) 23-39 (2001) SPB Academic Publishing bv, The Hague Hierarchical analysis of mtDNA variation and the use of mtDNA for isopod (Crustacea: Peracarida: Isopoda) systematics R. Wetzer Invertebrate Zoology, Crustacea, Natural History Museum of Los Angeles County, 900 Exposition Blvd., Los Angeles, CA 90007, USA, [email protected] Keywords: 12S rRNA, 16S rRNA, COI, mitochondrial DNA, isopod, Crustacea, molecular Abstract Transition/transversion bias 30 Discussion 32 Nucleotide composition 32 Carefully collected molecular data and rigorous analyses are Transition/transversionbias 34 revolutionizing today’s phylogenetic studies. Althoughmolecular Sequence divergences 34 data have been used to estimate various invertebrate phylogenies Rate variation across 34 lor lineages more than a decade,this is the first ofdifferent study survey Conclusions 35 of regions mitochondrial DNA in isopod crustaceans assessing Acknowledgements 36 sequence divergence and hence the usefulness ofthese regions References 36 to infer phylogeny at different hierarchical levels. 1 evaluate three loci fromthe mitochondrial ribosomal RNAs genome (two (12S, 16S) and oneprotein-coding (COI)) for their appropriateness in inferring isopod phylogeny at the suborder level and below. Introduction The patterns are similar for all three loci with the most speciose suborders ofisopods also having the most divergent mitochondrial The crustacean order Isopoda is important and nucleotide sequences. Recommendations for designing an or- because it has a broad dis- der- or interesting geographic suborder-level molecular study in previously unstudied groups of Crustacea tribution and is diverse. There would include: (1) collecting a minimum morphologically are of two-four species or genera thoughtto be most divergent, (2) more than 10,000 described marine, freshwater, sampling the across of interest as equally as possible in group and terrestrial species, ranging in length from 0.5 terms of taxonomic representation and the distributionofspecies, mm to 440 mm. -

Mountains, Streams, and Lakes of Oklahoma I
Information Series #1, June 1998 Mountains, Streams, and Lakes of OklahomaI Kenneth S. Johnson2 INTRODUCTION valleys, hills, and plains throughout most of the re mainder of Oklahoma (Fig. 1). All the major lakes and Mountains and streams define the landscape of reservoirs of Oklahoma are man-made, and they are Oklahoma (Fig. 1). The mountains consist mainly of important for flood contr()l, water supply, recreation, resistant rock masses that were folded, faulted, and and generation of hydroelectric power. Natural lakes thrust upward in the geologic past (Fig. 2), whereas in Oklahoma are limited to oxbow lakes along major the streams have persisted in eroding less-resistant streams and to playa lakes in the High Plains region rock units and lowering the landscape to form broad of the west. Alphabetical List of20 Lakes with Largest Surface Area (from Oklahoma Water Atlas, Oklahoma Water Resources Board) 1. Broken Bow 11. Lake 0' The Cherokees 2. Canton 12. Oologah 3. Eufaula 13. Robert s. Kerr 4. Fort Gibson 14. Sardis 5. Foss 15. Skiatook 6. Great Salt Plains 16. Tenkiller Ferry ·7. Hudson 17. Texoma 8. Hugo 18. Waurika 9. Kaw 19. Webbers Falls 10. Keystone 20. Wister Modified from Historical Atlas of Oklahoma, by John W. Morris, Charles R. Goins, and Edwin C. 25 McReynolds. Copyright © 1986 by the University I of Oklahoma Press. o 40 80Km Figure 1. Mountains, streams, and principal lakes of Oklahoma. lReprinted from Oklahoma Geology Notes (1993), vol. 53, no. 5, p. 180-188. The Notes article was reprinted and expanded from Oklahoma Almanac, 1993-1994, Oklahoma Department of Lihraries, p. -

Allocrangonyx Hubrichti)
Conservation Assessment for Hubricht’s Long-Tailed Amphipod (Allocrangonyx Hubrichti) (From Gardner, 1986) USDA Forest Service, Eastern Region October 2002 Julian J. Lewis, Ph.D. J. Lewis & Associates, Biological Consulting 217 W. Carter Avenue Clarksville, IN 47129 [email protected] MARK TWAIN NATIONAL FOREST This Conservation Assessment was prepared to compile the published and unpublished information on Allocrangonyx hubrichti. It does not represent a management decision by the U.S. Forest Service. Though the best scientific information available was used and subject experts were consulted in preparation of this document, it is expected that new information will arise. In the spirit of continuous learning and adaptive management, if you have information that will assist in conserving the subject community and associated taxa, please contact the Eastern Region of the Forest Service Threatened and Endangered Species Program at 310 Wisconsin Avenue, Milwaukee, Wisconsin 53203. Conservation Assessment for Hubricht’s Long-Tailed Amphipod (Allocrangonyx Hubrichti) 2 Table of Contents EXECUTIVE SUMMARY.....................................................................4 NOMENCLATURE AND TAXONOMY...............................................4 DESCRIPTION OF SPECIES................................................................4 LIFE HISTORY......................................................................................4 HABITAT ...............................................................................................5 DISTRIBUTION -

Federal Register / Vol. 62, No. 144 / Monday, July 28, 1997 / Proposed Rules 40319
Federal Register / Vol. 62, No. 144 / Monday, July 28, 1997 / Proposed Rules 40319 FEDERAL COMMUNICATIONS Species Act of 1973, as amended (Act), (egg-bearing) females have been COMMISSION for the Illinois cave amphipod, observed in summer and fall. Gammarus acherondytes, Historically, This species is best differentiated 47 CFR Chapter I the amphipod was known from six cave from other amphipods in the field, [CC Docket No. 97±146, FCC 97±219] streams in Monroe and St. Clair especially from G. fasciatus, which it counties, Illinois. Recent surveys have resembles, by its color, small degenerate Complete Detariffing for Competitive found the species at only three of the eyes, and a much longer first antenna. Access Providers and Competitive original six sites. This species is It is usually associated with the larger Local Exchange Carriers; Correction believed to be threatened primarily by G. troglophilus (Hubricht and Mackin groundwater pollution which is likely 1940) but is much less common AGENCY: Federal Communications due to the application of pesticides in (Holsinger 1972). Commission. cave stream recharge areas (the area of This species is a troglobitic (cave ACTION: Proposed rule; correction. land surface from which water drains dependent) species inhabiting the dark into a particular cave stream). A zone of cave streams. As a group, SUMMARY: The Federal Communications possible secondary threat is amphipods require cold water and are Commission published in the Federal contamination of subsurface water by intolerant of wide ranges in Register of July 17, 1997, a document human and animal wastes from sewage temperature. They are strongly sensitive concerning a Notice of Proposed and septic systems and livestock to touch and react negatively to light. -
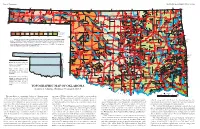
TOPOGRAPHIC MAP of OKLAHOMA Kenneth S
Page 2, Topographic EDUCATIONAL PUBLICATION 9: 2008 Contour lines (in feet) are generalized from U.S. Geological Survey topographic maps (scale, 1:250,000). Principal meridians and base lines (dotted black lines) are references for subdividing land into sections, townships, and ranges. Spot elevations ( feet) are given for select geographic features from detailed topographic maps (scale, 1:24,000). The geographic center of Oklahoma is just north of Oklahoma City. Dimensions of Oklahoma Distances: shown in miles (and kilometers), calculated by Myers and Vosburg (1964). Area: 69,919 square miles (181,090 square kilometers), or 44,748,000 acres (18,109,000 hectares). Geographic Center of Okla- homa: the point, just north of Oklahoma City, where you could “balance” the State, if it were completely flat (see topographic map). TOPOGRAPHIC MAP OF OKLAHOMA Kenneth S. Johnson, Oklahoma Geological Survey This map shows the topographic features of Oklahoma using tain ranges (Wichita, Arbuckle, and Ouachita) occur in southern contour lines, or lines of equal elevation above sea level. The high- Oklahoma, although mountainous and hilly areas exist in other parts est elevation (4,973 ft) in Oklahoma is on Black Mesa, in the north- of the State. The map on page 8 shows the geomorphic provinces The Ouachita (pronounced “Wa-she-tah”) Mountains in south- 2,568 ft, rising about 2,000 ft above the surrounding plains. The west corner of the Panhandle; the lowest elevation (287 ft) is where of Oklahoma and describes many of the geographic features men- eastern Oklahoma and western Arkansas is a curved belt of forested largest mountainous area in the region is the Sans Bois Mountains, Little River flows into Arkansas, near the southeast corner of the tioned below. -

Four Western Cheilanthoid Ferns in Oklahoma
Oklahoma Native Plant Record 65 Volume 10, December 2010 FOUR WESTERN CHEILANTHOID FERNS IN OKLAHOMA Bruce A. Smith McLoud High School McLoud, Oklahoma 74851 Keywords: arid, distribution, habitat, key ABSTRACT The diversity of ferns in some of the more arid climates of western Oklahoma is surprising. This article examines four Oklahoma cheilanthoid ferns: Astrolepis integerrima, Cheilanthes wootonii, Notholaena standleyi, and Pellaea wrightiana. With the exceptions of A. integerrima and P. wrightiana which occur in Alabama and North Carolina respectively, all four species reach their eastern limits of distribution in Oklahoma. Included in this article are common names, synonyms, brief descriptions, distinguishing characteristics, U.S. and Oklahoma distribution, habitat information, state abundance, and a dichotomous key to selected cheilanthoids. The Oklahoma Natural Heritage Inventory has determined that all but one (N. standleyi) are species of concern in the state. INTRODUCTION of eastern Oklahoma, while most members of the Pteridaceae occur in Almost half of the ferns in the family western Oklahoma (Taylor & Taylor Pteridaceae are xeric adapted ferns. In 1991). Oklahoma six genera and sixteen species Statewide, the most common species in the family are known to occur. They in the Pteridaceae is Pellaea atropurpurea live on dry or moist rocks and can be (Figure 4), which can be found found in rock crevices, at the bases of throughout the body of the state and boulders, or on rocky ledges. Common Cimarron County in the panhandle. The associated species include lichens, mosses, rarest are Cheilanthes horridula and liverworts, and spike mosses. Two Cheilanthes lindheimeri. Cheilanthes horridula physical characteristics that unite the and Cheilanthes lindheimeri have only been family are the marginal sori (Figure 1) and seen in one county each, Murray and the lack of a true indusium. -

The Biological Resources of Illinois Caves and Other
I LLINOI S UNIVERSITY OF ILLINOIS AT URBANA-CHAMPAIGN PRODUCTION NOTE University of Illinois at Urbana-Champaign Library Large-scale Digitization Project, 2007. EioD THE BIOLOGICAL RESOURCES OF ILLINOIS CAVES AND OTHER SUBTERRANEAN ENVIRONMENTS Determination of the Diversity, Distribution, and Status of the Subterranean Faunas of Illinois Caves and How These Faunas are Related to Groundwater Quality Donald W. Webb, Steven J. Taylor, and Jean K. Krejca Center for Biodiversity Illinois Natural History Survey 607 East Peabody Drive Champaign, Illinois 61820 (217) 333-6846 TECHNICAL REPORT 1993 (8) ILLINOIS NATURAL HISTORY SURVEY CENTER FOR BIODIVERSITY Prepared for: The Environmental Protection Trust Fund Commission and Illinois Department of Energy and Natural Resources Office of Research and Planning 325 W. Adams, Room 300 Springfield, IL 62704-1892 Jim Edgar, Governor John Moore, Director State of Illinois Illinois Department of Energy and Natural Resources ACKNOWLEDGEMENTS Funding for this project was provided through Grant EPTF23 from the Environmental Protection Trust Fund Commission, administered by the Department of Energy and Natural Resources (ENR). Our thanks to Doug Wagner and Harry Hendrickson (ENR) for their assistance. Other agencies that contributed financial support include the Shawnee National Forest (SNF) and the Illinois Department of Transportation (IDOT). Many thanks to Mike Spanel (SNF) and Rich Nowack (IDOT) for their assistance. Several agencies cooperated in other ways; we are. grateful to: Illinois Department of Conservation (IDOC); Joan Bade of the Monroe-Randolph Bi- County Health Department; Russell Graham and Jim Oliver, Illinois State Museum (ISM); Dr. J. E. McPherson, Zoology Department, Southern Illinois University at Carbondale (SIUC). Further contributions were made by the National Speleological Society, Little Egypt and Mark Twain Grottoes of the National Speleological Society, and the Missouri Speleological Survey. -

Description of the Coalgate Quadrangle
' *' DESCRIPTION OF THE COALGATE QUADRANGLE. By Joseph A. Taff. GEOGRAPHY. forms of the Ouachita Mountain Range. The a long time the valleys become wide and silted, that at ordinary conditions the stream meanders ridges of the valley region are nearly parallel and the hills are gradually reduced to the level in rivulets or narrow channels. Indeed, its chan GENERAL RELATIONS. with those of the range, but, with the exception of the valleys. nel is so choked with sand that the water does The Coalgate quadrangle is bounded by the of the few isolated mountains which lie in Arkan The surface of the Coalgate quadrangle is of not at any stage of the river flow on the country meridians 96° and 96° 30' and the parallels 34° sas Valley, they have low relief. low relief, and the topographic features indicate rock. During floods, which usually come in 30' and 35°, and thus occupies one- Extentand The Prairie Plains region stretches from the that it has been so for a relatively long period of spring from the headwaters of the river, vast quarter of a square degree of the earth's th1?Juad=f Arkansas Valley and Ozark highland regions time. The larger streams have nearly ceased cut quantities of sands are swept down, shifting for surface. It is 34.4 miles long north rangle> northward and westward across north- Character ting their valleys deeper, and throughout most of mer shoals and channels. Little River, which is and south and 28.5 miles wide, and contains west Indian Territory into Oklahoma their courses are meandering in the deposits of silt tributary to the Canadian, crosses the northwest nearly 980 square miles. -

Ouachita Mountains Ecoregional Assessment December 2003
Ouachita Mountains Ecoregional Assessment December 2003 Ouachita Ecoregional Assessment Team Arkansas Field Office 601 North University Ave. Little Rock, AR 72205 Oklahoma Field Office 2727 East 21st Street Tulsa, OK 74114 Ouachita Mountains Ecoregional Assessment ii 12/2003 Table of Contents Ouachita Mountains Ecoregional Assessment............................................................................................................................i Table of Contents ........................................................................................................................................................................iii EXECUTIVE SUMMARY..............................................................................................................1 INTRODUCTION..........................................................................................................................3 BACKGROUND ...........................................................................................................................4 Ecoregional Boundary Delineation.............................................................................................................................................4 Geology..........................................................................................................................................................................................5 Soils................................................................................................................................................................................................6 -

Illinois Cave Amphipod (Gammarus Acherondytes) Recovery Plan
Illinois Cave Amphipod (Gammarus acherondytes) Recovery Plan U.S. Fish and Wildlife Service Great Lakes-Big Rivers Region (Region 3) Fort Snelling, Minnesota llIinois Cave Amphipod ( Gammarus acherondytes) Recovery Plan Prepared by Illinois Cave Amphipod Recovery Team Ms. Joan Bade, Recovery Team Leader Dr. Julian J. Lewis Illinois Department of Natural Resources J. Lewis and Associates BioI. Consulting Chester, Illinois Clarksville, Indiana Dr. Steven J. Taylor Ms. Diane Tecic Illinois Natural History Survey Illinois Department of Natural Resources Champaign, Illinois Granite City, Illinois Dr. Donald W. Webb Mr. Dennis Brand Illinois Natural History Survey Waterloo, Illinois Champaign, Illinois Mr. Kenneth Hartman, Jr. Dr. Samuel V. Panno Waterloo, Illinois Illinois State Geological Survey Champaign, Illinois Mr. Philip Moss Ozark Underground Laboratory Protem, Missouri For , Region 3 U.S. Fish and Wildlife Service Ft. Snelling, Minnesota 55111-4056 ~ Appro~e~:, (;)UQ l ~ - f'"J-"\ Regional Director, U,S. Fish and Wildlir.JService Date: j l2.4.1l () 'L Disclaimer Recovery plans delineate reasonable actions which are believed to be required to recover and/or protect listed species. Plans are published by the U.S. Fish and Wildlife Service (USFWS), sometimes prepared with the assistance of recovery teams, contractors, State agencies, and others. Objectives will be attained and any necessary funds made available subject to budgetary and other constraints affecting the parties involved, as well as the need to address other priorities. Recovery plans do not necessarily represent the views nor the official positions or approval of any individuals or agencies involved in the plan formulation, other than the USFWS. They represent the official position of the USFWS only after they have been signed by the Regional Director or Director as approved. -

Age of the Folding of the Oklahoma Mountains— % the Ouachita, Arbuckle, and Wichita Moun Tains of Oklahoma and the Llano-Burnet and Marathon Uplifts of Texas 1
BULLETIN OF THE GEOLOGICAL SOCIETY OF AMERICA Vol. 39. pp. 1031-1072 December 30. 1928 AGE OF THE FOLDING OF THE OKLAHOMA MOUNTAINS— % THE OUACHITA, ARBUCKLE, AND WICHITA MOUN TAINS OF OKLAHOMA AND THE LLANO-BURNET AND MARATHON UPLIFTS OF TEXAS 1 BY SIDNEY POWERS 2 (Presented by title before the Society December SI, 1927) CONTENTS Tage Introduction ............................................................................................................. 1031 Stratigraphy ............................................................................................................. 1036 The, Ouachita Mountains ..................................................................................... 1037 Problems............................................................................................................. 1037 Age of the Carboniferous sediments .......................................................... 1038 Origin of the “glacial” boulders .................................................................. 1042 Date of the folding and overthrusting ...................................................... 1047 The Arbuckle Mountains ........................................................................................ 1049 The Wichita Mountains ........................................................................................ 1056 Former extent ................................................................................................. 1056 Criner Hills ....................................................................................................