Full Text in Pdf Format
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Portugal's Hidden
The Azores Portugal ’s hidden gem AUGUST 5-13, 2021 $ The 400 COUPLE SavePER Book by Azores JANUARY 31, 2021 Dear Vanderbilt Traveler, Portugal ’s hidden gem You are invited to discover the exotic natural wonders of the Azores, Portugal’s hidden gem! We are pleased to announce a very special alumni trip to this cutting-edge travel destination scheduled for August 2021. One of two autonomous regions of Portugal, this archipelago is composed of nine volcanic islands in the Macaronesia region of the North Atlantic Ocean. Situated 930 miles directly west of Lisbon, this lush and untamed archipelago offers visitors an unexplored paradise replete with natural and culinary treasures. On the island of Pico, see the beauty of Arcos do Cachorro, Mistério de São João, and Fajã dos Vimes. Learn about the island’s fascinating natural history at Gruta das Torres, the largest lava tube in the Azores. Continue to the island of São Miguel and experience its breathtaking beauty at Sete Cidades, Terra Nostra Park, Fogo Lake, and the hot springs of Furnas. As stunning as the Azores geography can be, their cuisine is equally rewarding. Please your palette during a private visit to a local winery and cheese factory in Ponta dos Rosais. From the famous cozido to the local pastries and tea plantations, savor the wide range of Azorean tastes and traditions. Space on this program is strictly limited. Contact us as soon as possible to reserve your place. We look forward to having you join us on this remarkable adventure. Sincerely, Cary DeWitt Allyn For more details: WWW.VUCONNECT.COM/TRAVEL Director, Vanderbilt Travel Program 615.322.3673 AUGUST 5-13, 2021 THURSDAY, AUGUST 5: DEPART USA ALDEIA DA FONTE NATURE HOTEL Depart USA on your overnight flight(s) to Ponta Delgada, Azores. -

Checklists of Crustacea Decapoda from the Canary and Cape Verde Islands, with an Assessment of Macaronesian and Cape Verde Biogeographic Marine Ecoregions
Zootaxa 4413 (3): 401–448 ISSN 1175-5326 (print edition) http://www.mapress.com/j/zt/ Article ZOOTAXA Copyright © 2018 Magnolia Press ISSN 1175-5334 (online edition) https://doi.org/10.11646/zootaxa.4413.3.1 http://zoobank.org/urn:lsid:zoobank.org:pub:2DF9255A-7C42-42DA-9F48-2BAA6DCEED7E Checklists of Crustacea Decapoda from the Canary and Cape Verde Islands, with an assessment of Macaronesian and Cape Verde biogeographic marine ecoregions JOSÉ A. GONZÁLEZ University of Las Palmas de Gran Canaria, i-UNAT, Campus de Tafira, 35017 Las Palmas de Gran Canaria, Spain. E-mail: [email protected]. ORCID iD: 0000-0001-8584-6731. Abstract The complete list of Canarian marine decapods (last update by González & Quiles 2003, popular book) currently com- prises 374 species/subspecies, grouped in 198 genera and 82 families; whereas the Cape Verdean marine decapods (now fully listed for the first time) are represented by 343 species/subspecies with 201 genera and 80 families. Due to changing environmental conditions, in the last decades many subtropical/tropical taxa have reached the coasts of the Canary Islands. Comparing the carcinofaunal composition and their biogeographic components between the Canary and Cape Verde ar- chipelagos would aid in: validating the appropriateness in separating both archipelagos into different ecoregions (Spalding et al. 2007), and understanding faunal movements between areas of benthic habitat. The consistency of both ecoregions is here compared and validated by assembling their decapod crustacean checklists, analysing their taxa composition, gath- ering their bathymetric data, and comparing their biogeographic patterns. Four main evidences (i.e. different taxa; diver- gent taxa composition; different composition of biogeographic patterns; different endemicity rates) support that separation, especially in coastal benthic decapods; and these parametres combined would be used as a valuable tool at comparing biotas from oceanic archipelagos. -

Post-Glacial Filling of a Semi-Enclosed Basin: the Arguin Basin (Mauritania)
Marine Geology 349 (2014) 126–135 Contents lists available at ScienceDirect Marine Geology journal homepage: www.elsevier.com/locate/margeo Post-glacial filling of a semi-enclosed basin: The Arguin Basin (Mauritania) N. Aleman a,⁎,R.Certaina,J.P.Barusseaua,T.Courpa,A.Diab a Centre Européen de Formation et de Recherche sur les Environnements Méditerranéens, UMR5110, Université de Perpignan, 52 av. P Alduy, 66860 Perpignan, France b Institut Mauritanienne de Recherche Océanographique et des Pêches, BP22, Nouadhibou, Mauritania article info abstract Article history: Semi-enclosed basins are not very common features in the world and are most frequently the result of tectonic Received 31 January 2013 movements. Studies of their filling are usually based on the micropaleontological analyses of sediment cores Received in revised form 11 December 2013 (Torgersen et al., 1988; Reeves et al., 2007) or seismic analyses (Lykousis et al., 2007; Çagatay et al., 2009; Van Accepted 24 December 2013 Daele et al., 2011). The morphology of semi-enclosed basins is generally simple and bowl-shaped, and their Available online 2 January 2014 edges are marked by one or more sills. Their depths range from a few dozen to several thousand meters. Semi- Communicated by J.T. Wells enclosed basins are however present in some regions in the world. The semi-enclosed basin of the Golfe d'Arguin (Northwest Africa) is present on a wide, shallow shelf, bordering the Sahara desert, in a stable tectonic context. Its Keywords: sedimentary filling took place during the end of the post-glacial transgression. The current knowledge on sedi- semi-enclosed basin mentary filling of semi-enclosed basins is rather limited and inadequate to fully understand the processes at play. -

The Best of the Azores
The Best of the Azores 11 October to 21 October, 2017 $2,799 Per Person…based on double occupancy. Includes taxes of approximately 160 USD. 9 Nights with breakfast daily in Terceira, Faial, and Sao Miguel. 16 Meals, including welcome and farewell dinner. Private Tours: Terceira Island Tour, Faial Island Tour, Pico Island Tour with ferry, Sete Cidades Tour, Furnas Tour (with Cozido) ,all private tours include transfers and English Speaking Guide. Airfare included from Boston to Terceira, return from Sao Miguel to Boston as well as internal flights within the Azores. Includes Round-Trip Transfer from New Bedford to Boston Logan Airport. 761 Bedford Street, Fall River, MA 02723 www.sagresvacations.com Ph#508-679-0053 Your Itinerary Includes Hotels Angra do Heroismo, Terceira Island o Angra Garden Hotel Check in 12OCT-14OCT o Double Room with breakfast daily Horta, Faial Island o Faial Resort Hotel Check in 14OCT-16OCT o Double room with breakfast daily Ponta Delgada, Sao Miguel Island o Royal Garden Hotel Check in 16OCT-21OCT Double Room with breakfast daily Private Transfers Airport Transfers Included o New Bedford to Boston Logan Airport transfers round-trip 11OCT-21 OCT o TER Airport to Angra Garden Hotel and vice-versa 12OCT-14OCT o Horta Airport to Faial Resort Hotel and vice-versa 14OCT-16OCT. o P. Delgada Airport to Royal Garden Hotel and vice-versa Terceira 16OCT-21OCT. All Transfers included on Private Tours Private Tours/Excursions Terceira o Full Day Terceira Tour with Lunch. Faial o Full Day Faial Tour with Lunch o Pico -

Secret Base: Santa Maria Airfield During World War II
vol.3 | Dezembro 2017 | Revista LPAZ vol.3 | Dezembro 2017 | Revista LPAZ September. At first base personnel were mostly civilian activated on August 1, 1944, with assignment to the North Secret Base: workers from two construction companies: W. C. Shepard Atlantic Division of the Air Transport Command. of Atlanta, Georgia, and W. A. Hart of Fort Lauderdale, Santa Maria Airfield during World War II Florida. Records show that during the opening phase, Keeping the military nature of the base secret was difficult. there were 267 civilians, 11 officers, and 124 enlisted Mail and baggage was inspected, with strict censorship of Daniel Haulman men, about half of them members of the 786th Military as many as 800 letters a day. Portuguese natives were Air Force Historical Research Agency | USA Police Detachment. Lt. Col. Russell W. Gray commanded not fooled when they saw American military personnel the temporary camp, with 45 pyramidal tents for military wearing khaki shirts showing marks where chevrons and During World War II, many European nations remained occupied France. In August, free French and American personnel. Nine other tents sheltered administrative insignia had been. Many of the American civilian workers neutral in the struggle between the Axis countries, led by forces invaded southern France, where German forces offices, mess facilities, and supplies. Civilian workers became discipline problems, partly because they had not Germany, Italy, and Japan, and the Allies, led by Britain, were forced to retreat, and Allied forces entered Paris by lived in similar temporary shelters, at first. By the end been carefully selected, and partly because they were the Soviet Union, and the United States. -

Distributional Impacts of Fisheries Subsidies and Their Reform Case Studies of Senegal and Vietnam
Distributional impacts of fisheries subsidies and their reform Case studies of Senegal and Vietnam Sarah Harper and U Rashid Sumaila Working Paper Fisheries; Sustainable markets Keywords: March 2019 Sustainable fisheries; fossil fuel subsidies; livelihoods; gender and generation; equity About the authors Sarah Harper, Fisheries Economics Research Unit, Institute for the Oceans and Fisheries, University of British Columbia; U Rashid Sumaila, Fisheries Economics Research Unit, Institute for the Oceans and Fisheries, University of British Columbia Corresponding author email [email protected] Produced by IIED’s Shaping Sustainable Markets Group The Shaping Sustainable Markets group works to make sure that local and global markets are fair and can help poor people and nature to thrive. Our research focuses on the mechanisms, structures and policies that lead to sustainable and inclusive economies. Our strength is in finding locally appropriate solutions to complex global and national problems. Partner organisation The Fisheries Economics Research Unit strives for interdisciplinary solutions to global, national, and local marine and freshwater management issues. We focus on economic and policy analysis and mobilize researchers, students, and practitioners to advance resource management for the benefit of current and future generations, while maintaining ‘healthy’ ecosystems. Published by IIED, March 2019 Sarah Harper, U Rashid Sumaila (2019) Distributional impacts of fisheries subsidies and their reform: case studies of Senegal and Vietnam. -

GPS AZORES Project
GPS AZORES Project Title: Geopolitical framework of the Macaronesia region Project Coordinator: Helena Calado (Universidade dos Azores) Work Package Coordinator: Juan Luis Suarez de Vivero (Universidad de Sevilla) Authors: Elisabetta Menini, Firdaous Halim, Daniela Gabriel, Juan Luis Suarez de Vivero, Helena Calado, Fabiana Moniz, Mario Caña Varona. Submission date: 31 August 2018 Acknowledgements: Christine Ladiero for helping with statistics Citation: Menini E., Halim F., Gabriel, D., Suarez de Vivero, JL., Calado, H., Moniz, F., Caña Varona, M. 2018. Geopolitical framework of the Macaronesia region. GPS Azores project: Ponta Delgada. Page 1 This project was financed in 85% by FEDER and in 15% with regional funds through the Programa Operacional Açores 2020 (Operational Program Azores 2020), in scope of the project « GPSAZORES -ACORES-01-0145-FEDER-00002». Page 2 Contents ACRONYMS 4 1. Maritime scenario characterisation: Main political, jurisdictional and socio-economic features 5 1.1 Political geography of the region 5 1.1.1 The regional context: Macaronesia 5 1.1.2 Countries and territories 12 1.2 Maritime space 16 1.2.1 Macaronesia in the context of the United Nations Convention of the Law of the Sea 16 1.2.2 Maritime jurisdictions 19 1.2.3 Maritime borders 21 1.3 The socio-economic context: exploitation and uses of the maritime space 26 1.3.1 Demography 26 1.3.2 Economic development 27 1.3.3 Tourism 29 1.3.1 Fisheries and aquaculture 31 1.3.2 Regional maritime geo-economics 33 1.3.3 Other relevant maritime economic activities 33 -

The Age of Exploration (Also Called the Age of Discovery) Began in the 1400S and Continued Through the 1600S. It Was a Period Of
Activity 1 of 3 for NTI May 18 - 22 - Introduction to Exploration of North America Go to: https://www.ducksters.com/history/renaissance/age_of_exploration_and_discovery.php Click on the link above to read the article. There is a feature at the bottom that will allow you to have the text read to you, if you want. After you read the article, answer the questions below. You can highlight or bold your answers if completing electronically. I have copied the website text below if you need it. The Age of Exploration (also called the Age of Discovery) began in the 1400s and continued through the 1600s. It was a period of time when the European nations began exploring the world. They discovered new routes to India, much of the Far East, and the Americas. The Age of Exploration took place at the same time as the Renaissance. Why explore? Outfitting an expedition could be expensive and risky. Many ships never returned. So why did the Europeans want to explore? The simple answer is money. Although, some individual explorers wanted to gain fame or experience adventure, the main purpose of an expedition was to make money. How did expeditions make money? Expeditions made money primarily by discovering new trade routes for their nations. When the Ottoman Empire captured Constantinople in 1453, many existing trade routes to India and China were shut down. These trade routes were very valuable as they brought in expensive products such as spices and silk. New expeditions tried to discover oceangoing routes to India and the Far East. Some expeditions became rich by discovering gold and silver, such as the expeditions of the Spanish to the Americas. -

Remembering Dr. Kernohan: "She Enjoyed Helping People"
I Perspectives Remembering Dr. Kernohan: "She enjoyed helping people" Barbara Yaffe r. Mary Kernohan who chose a career that allowed fore she came home from work; D used to fly out of her to do that in an intimate way. her car keys and some money Nain and Goose Bay Mary Kernohan was mur- were stolen and her pick-up truck in a single-engine dered last fall. She was 32. Ac- was driven to another part of plane to deliver medical care to cording to Sgt. Joseph Macdonald town, where it was discovered by people living in remote villages of the Royal Canadian Mounted police. on Labrador's rugged coast. Police (RCMP) in Goose Bay, she Court proceedings are pend- Sometimes weather would ex- was beaten "viciously" with a ing to review the provincial tend a brief visit to a week or baseball bat on Nov. 13 and died court's January decision to try the more and snow drifts would be 8 days later. The murder left boy in adult court. If he is tried so severe she would have to many people dazed - residents and convicted in youth court, the crawl out a second-storey win- of Happy Valley/Goose Bay, her maximum sentence would be 3 dow in order to leave her nurs- family in Nova Scotia and her years' imprisonment; in adult ing-station office. fiance and colleague, Dr. David court, he could face life imprison- She was a woman with a Beach. ment. sense of fun and adventure and Just 2 hours after a neigh- Neither Beach nor the Ker- strong convictions about public bour found her, lying bloodied in nohan family is too preoccupied service, a high school valedictori- the kitchen of her Happy Valley with the upcoming court pro- an who liked helping people and home, a 16-year-old boy was ar- ceedings or with the fate of the rested and charged with second- murderer, beyond wanting the Barbara Yaffe is a freelance writer living degree murder. -
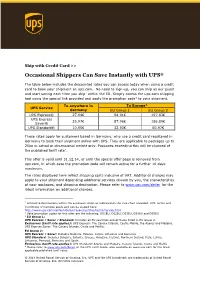
Occasional Shippers Can Save Instantly with UPS®
Ship with Credit Card >> Occasional Shippers Can Save Instantly with UPS® The table below includes the discounted rates you can access today when using a credit card to book your shipment on ups.com. No need to sign-up, you can ship as our guest and start saving each time you ship1 within the EU. Simply access the ups.com shipping tool using the special link provided and apply the promotion code2 to your shipment. To anywhere in To Europe3 UPS Service Germany EU Group 1 EU Group 2 UPS Express® 27.94€ 94.01€ 197.83€ UPS Express 25.97€ 87.96€ 186.89€ Saver® UPS Standard® 10.95€ 32.93€ 50.97€ These rates apply for customers based in Germany, who use a credit card registered in Germany to book their shipment online with UPS. They are applicable to packages up to 20kg in actual or dimensional weight only. Packages exceeding this will be charged at the published tariff rate4. This offer is valid until 31.12.14, or until the special offer page is removed from ups.com, in which case the promotion code will remain active for a further 10 days maximum. The rates displayed here reflect shipping costs inclusive of VAT. Additional charges may apply to your shipment depending additional services chosen by you, the characteristics of your packages, and shipping destination. Please refer to www.ups.com/de/en for the latest information on additional charges. 1 Limited to destinations within the European Union as indicated on the rate chart provided. UPS Terms and Conditions of Carriage apply and can be viewed here: http://www.ups.com/content/de/en/resources/ship/terms/service.html 2 Valid promotion codes for this offer are the following: DE1EU, DE2EU, DE3EU, DE4EU and DE5EU 3 EU Group 1: UPS Express / Saver / Standard: Includes all EU countries except those listed in EU Group 2. -

Santa Maria Island's (Azores Archipelago) Geological Evolution: New Insights from High-Resolution Marine Geophysical Data
Geophysical Research Abstracts Vol. 20, EGU2018-2422, 2018 EGU General Assembly 2018 © Author(s) 2017. CC Attribution 4.0 license. Santa Maria Island’s (Azores Archipelago) geological evolution: new insights from high-resolution marine geophysical data. Alessandro Ricchi (1), Rui Quartau (2,3), Ricardo Ramalho (4,5), Claudia Romagnoli (1), Daniele Casalbore (6,7) (1) University of Bologna, BiGeA, Italy ([email protected]), (2) Instituto Dom Luiz, Faculdade de Ciências, Universidade de Lisboa, 1749-016 Lisboa, Portugal, (3) Divisão de Geologia Marinha, Instituto Hidrográfico – Marinha, 1200-615 Lisboa, Portugal, (4) School of Earth Sciences, University of Bristol, BS8 1RJ Bristol, UK, (5) Lamont-Doherty Earth Observatory at Columbia University, 10964 New York, USA, (6) Sapienza Università di Roma, Dipartimento Scienze della Terra, Roma, Italy, (7) Istituto di Geologia Ambientale e Geoingegneria, Consiglio Nazionale delle Ricerche, Area della Ricerca Roma 1, Montelibretti, Via Salaria Km 29,300, Monterotondo, Roma, Italy Santa Maria is the oldest island of the Azores Archipelago in the Atlantic Ocean, having first emerged at c.6 Ma (Ramalho et al., 2017). Its geological evolution up to now was constrained only by the knowledge of the subaerial volcanic edifice, since the submerged portion of the island has never been studied. In this work we used high-resolution multibeam bathymetry and closely-spaced seismic reflection profiles to study the morpho-stratigraphic setting of Santa Maria’s insular shelf. In particular the shelf width and the depth of its edge where used as proxies for reconstructing the development of the earlier portions of Santa Maria Island. Our study suggests that the large insular shelf off the northern coast of Santa Maria is the remnant of an early, now almost completely razed, volcanic edifice likely older than the volcanic sequences exposed onland. -

Housing Demand and Supply in Central Labrador: Housing in the Happy Valley- Goose Bay Cluster
HOUSING DEMAND AND SUPPLY IN CENTRAL LABRADOR: HOUSING IN THE HAPPY VALLEY- GOOSE BAY CLUSTER STEPHEN B. JEWCZYK JULY 2018 REPORT PREPARED FOR THE LESLIE HARRIS CENTRE OF REGIONAL POLICY AND DEVELOPMENT, MEMORIAL UNIVERSITY 1 POPULATION PROJECT: NEWFOUNDLAND AND LABRADOR IN TRANSITION Housing Demand and Supply in Central Labrador HOUSING IN THE HAPPY VALLEY-GOOSE BAY CLUSTER REPORT PREPARED FOR THE LESLIE HARRIS CENTRE OF REGIONAL POLICY AND DEVELOPMENT, MEMORIAL UNIVERSITY POPULATION PROJECT: NEWFOUNDLAND AND LABRADOR IN TRANSITION July 2018 Stephen B. Jewczyk. B.A.A., (Urban and Regional Planning), FCIP1 1 I extend my sincere thanks to all individuals who were interviewed as part of this study and attended the community consultations. I also thank Keith Storey for his valuable comments and editorial contributions. The Population Project: Newfoundland and Labrador in Transition In 2015, Newfoundland and Labrador had the most rapidly aging population in the country – which when combined with high rates of youth out-migration, declining birth rates, and an increasing number of people moving from rural parts of the province to more urban centres, means that the province is facing an unprecedented population challenge. Without intervention, this trend will have a drastic impact on the economy, governance, and the overall quality of life for the people of the province. Planning for this change and developing strategies to adjust and adapt to it is paramount. The Harris Centre’s Population Project has developed potential demographic scenarios