The Induction Levels of Heat Shock Protein 70 Differentiate the Vulnerabilities to Mutant Huntingtin Among Neuronal Subtypes
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

The HECT Domain Ubiquitin Ligase HUWE1 Targets Unassembled Soluble Proteins for Degradation
OPEN Citation: Cell Discovery (2016) 2, 16040; doi:10.1038/celldisc.2016.40 ARTICLE www.nature.com/celldisc The HECT domain ubiquitin ligase HUWE1 targets unassembled soluble proteins for degradation Yue Xu1, D Eric Anderson2, Yihong Ye1 1Laboratory of Molecular Biology, National Institute of Diabetes and Digestive and Kidney Diseases, National Institutes of Health, Bethesda, MD, USA; 2Advanced Mass Spectrometry Core Facility, National Institute of Diabetes and Digestive and Kidney Diseases, National Institutes of Health, Bethesda, MD, USA In eukaryotes, many proteins function in multi-subunit complexes that require proper assembly. To maintain complex stoichiometry, cells use the endoplasmic reticulum-associated degradation system to degrade unassembled membrane subunits, but how unassembled soluble proteins are eliminated is undefined. Here we show that degradation of unassembled soluble proteins (referred to as unassembled soluble protein degradation, USPD) requires the ubiquitin selective chaperone p97, its co-factor nuclear protein localization protein 4 (Npl4), and the proteasome. At the ubiquitin ligase level, the previously identified protein quality control ligase UBR1 (ubiquitin protein ligase E3 component n-recognin 1) and the related enzymes only process a subset of unassembled soluble proteins. We identify the homologous to the E6-AP carboxyl terminus (homologous to the E6-AP carboxyl terminus) domain-containing protein HUWE1 as a ubiquitin ligase for substrates bearing unshielded, hydrophobic segments. We used a stable isotope labeling with amino acids-based proteomic approach to identify endogenous HUWE1 substrates. Interestingly, many HUWE1 substrates form multi-protein com- plexes that function in the nucleus although HUWE1 itself is cytoplasmically localized. Inhibition of nuclear entry enhances HUWE1-mediated ubiquitination and degradation, suggesting that USPD occurs primarily in the cytoplasm. -

Heat Shock Protein 27 Inhibits HMGB1 Translocation by Regulating CBP
Molecular Immunology 108 (2019) 45–55 Contents lists available at ScienceDirect Molecular Immunology journal homepage: www.elsevier.com/locate/molimm Heat shock protein 27 inhibits HMGB1 translocation by regulating CBP acetyltransferase activity and ubiquitination T ⁎⁎ Xiaowen Bia, Miao Xua, Jinfei Lia, Ting Huanga, Baolin Jianga, Lei Shena, Lan Luob, , ⁎⁎⁎ ⁎ Shixiang Liuc, , Zhimin Yina, a Jiangsu Province Key Laboratory for Molecular and Medical Biotechnology, College of Life Science, Nanjing Normal University, Nanjing, Jiangsu, PR China b State Key Laboratory of Pharmaceutical Biotechnology, School of Life Sciences, Nanjing University, Nanjing, Jiangsu, PR China c Jurong People’s Hospital, Zhenjiang, Jiangsu, PR China ARTICLE INFO ABSTRACT Keywords: Heat-shock protein 27 (Hsp27) is a member of the small heat shock protein family that has been reported to Hsp27 protect cells against pro-inflammatory stresses. High mobility group box 1 (HMGB1) is a proinflammatory cy- CBP tokine associated with death from sepsis and other inflammatory diseases. After being acetylated by CREB- HMGB1 binding protein (CBP), the transcriptional adaptor and acetyltransferase, HMGB1 translocates from the nucleus Phosphorylation to the cytoplasm. In the present study, we investigated the effects of Hsp27 on HMGB1 translocation from the Acetylation nucleus to the cytoplasm in THP-1 cells. We found that Hsp27 phosphorylation decreased LPS-induced HMGB1 acetylation and translocation from the nucleus to the cytoplasm, as well as its release from THP-1 cells. The study further showed that cytosolic non-phosphorylated Hsp27 enhanced CBP ubiquitination and degradation in LPS-unstimulated cells, which suggested that Hsp27 maintained suitable CBP levels under normal physiological conditions. After LPS stimulation, Hsp27 was phosphorylated at serine residues 15/78 and translocated from the cytoplasm into the nucleus. -

Runx1 Gene in Acute Myeloid Leukemia: Expression Level Signifcance and Impact on Clinical Features
Runx1 Gene in Acute Myeloid Leukemia: Expression Level Signicance and Impact on Clinical Features Fadwa Said Abdelazim Mohamed ( [email protected] ) Cairo University Roxan E Shak National Cancer Institute, Cairo University Naglaa M. Hassan National Cancer Institute, Cairo University Research article Keywords: Acute myeloid leukemia, RUNX1, mutation, gene expression, Egypt Posted Date: April 7th, 2020 DOI: https://doi.org/10.21203/rs.3.rs-20926/v1 License: This work is licensed under a Creative Commons Attribution 4.0 International License. Read Full License Page 1/20 Abstract Background Acute myeloid leukemia represents the highest percentage of all adult acute leukemia variants. Runt-related transcription factor1 (RUNX1), a transcription factor with a known tumor suppressor function was recently reported as a tumor promotor in AML. We investigated the role of RUNX1 geneexpression levelin Egyptian AML patients and delineateits clinical signicance. Methods This study recruited 91 AML patients that were recently diagnosed at our hospital with 14 healthy age- and sex-matched donors of bone marrow transplantation unit. We measured RUNX1 gene expression level using reverse transcription–quantitative polymerase chain reaction. Results We found that RUNX1 gene expression level was signicantly higher compared to the control group (p < 0.001). Patients with FLT3 mutations had the higher expression level of RUNX1 (p=0.023). The male patients expressed signicantly higher level of RUNX1 (p=0.046). Conclusion The RUNX1 gene expression may serve as a diagnostic marker in Egyptian AML patients. Its relation to FLT3 may give clue that patients carrying this mutation may benet fromnew treatments that target RUNX1 in the future. -

Genome-Wide Sirna Screen for Mediators of NF-Κb Activation
Genome-wide siRNA screen for mediators SEE COMMENTARY of NF-κB activation Benjamin E. Gewurza, Fadi Towficb,c,1, Jessica C. Marb,d,1, Nicholas P. Shinnersa,1, Kaoru Takasakia, Bo Zhaoa, Ellen D. Cahir-McFarlanda, John Quackenbushe, Ramnik J. Xavierb,c, and Elliott Kieffa,2 aDepartment of Medicine and Microbiology and Molecular Genetics, Channing Laboratory, Brigham and Women’s Hospital and Harvard Medical School, Boston, MA 02115; bCenter for Computational and Integrative Biology, Massachusetts General Hospital, Harvard Medical School, Boston, MA 02114; cProgram in Medical and Population Genetics, The Broad Institute of Massachusetts Institute of Technology and Harvard, Cambridge, MA 02142; dDepartment of Biostatistics, Harvard School of Public Health, Boston, MA 02115; and eDepartment of Biostatistics and Computational Biology and Department of Cancer Biology, Dana-Farber Cancer Institute, Boston, MA 02115 Contributed by Elliott Kieff, December 16, 2011 (sent for review October 2, 2011) Although canonical NFκB is frequently critical for cell proliferation, (RIPK1). TRADD engages TNFR-associated factor 2 (TRAF2), survival, or differentiation, NFκB hyperactivation can cause malig- which recruits the ubiquitin (Ub) E2 ligase UBC5 and the E3 nant, inflammatory, or autoimmune disorders. Despite intensive ligases cIAP1 and cIAP2. CIAP1/2 polyubiquitinate RIPK1 and study, mammalian NFκB pathway loss-of-function RNAi analyses TRAF2, which recruit and activate the K63-Ub binding proteins have been limited to specific protein classes. We therefore under- TAB1, TAB2, and TAB3, as well as their associated kinase took a human genome-wide siRNA screen for novel NFκB activa- MAP3K7 (TAK1). TAK1 in turn phosphorylates IKKβ activa- tion pathway components. Using an Epstein Barr virus latent tion loop serines to promote IKK activity (4). -

At Elevated Temperatures, Heat Shock Protein Genes Show Altered Ratios Of
EXPERIMENTAL AND THERAPEUTIC MEDICINE 22: 900, 2021 At elevated temperatures, heat shock protein genes show altered ratios of different RNAs and expression of new RNAs, including several novel HSPB1 mRNAs encoding HSP27 protein isoforms XIA GAO1,2, KEYIN ZHANG1,2, HAIYAN ZHOU3, LUCAS ZELLMER4, CHENGFU YUAN5, HAI HUANG6 and DEZHONG JOSHUA LIAO2,6 1Department of Pathology, Guizhou Medical University Hospital; 2Key Lab of Endemic and Ethnic Diseases of The Ministry of Education of China in Guizhou Medical University; 3Clinical Research Center, Guizhou Medical University Hospital, Guiyang, Guizhou 550004, P.R. China; 4Masonic Cancer Center, University of Minnesota, Minneapolis, MN 55455, USA; 5Department of Biochemistry, China Three Gorges University, Yichang, Hubei 443002; 6Center for Clinical Laboratories, Guizhou Medical University Hospital, Guiyang, Guizhou 550004, P.R. China Received December 16, 2020; Accepted May 10, 2021 DOI: 10.3892/etm.2021.10332 Abstract. Heat shock proteins (HSP) serve as chaperones genes may engender multiple protein isoforms. These results to maintain the physiological conformation and function of collectively suggested that, besides increasing their expres‑ numerous cellular proteins when the ambient temperature is sion, certain HSP and associated genes also use alternative increased. To determine how accurate the general assumption transcription start sites to produce multiple RNA transcripts that HSP gene expression is increased in febrile situations is, and use alternative splicing of a transcript to produce multiple the RNA levels of the HSF1 (heat shock transcription factor 1) mature RNAs, as important mechanisms for responding to an gene and certain HSP genes were determined in three cell increased ambient temperature in vitro. lines cultured at 37˚C or 39˚C for three days. -

The UBE2L3 Ubiquitin Conjugating Enzyme: Interplay with Inflammasome Signalling and Bacterial Ubiquitin Ligases
The UBE2L3 ubiquitin conjugating enzyme: interplay with inflammasome signalling and bacterial ubiquitin ligases Matthew James George Eldridge 2018 Imperial College London Department of Medicine Submitted to Imperial College London for the degree of Doctor of Philosophy 1 Abstract Inflammasome-controlled immune responses such as IL-1β release and pyroptosis play key roles in antimicrobial immunity and are heavily implicated in multiple hereditary autoimmune diseases. Despite extensive knowledge of the mechanisms regulating inflammasome activation, many downstream responses remain poorly understood or uncharacterised. The cysteine protease caspase-1 is the executor of inflammasome responses, therefore identifying and characterising its substrates is vital for better understanding of inflammasome-mediated effector mechanisms. Using unbiased proteomics, the Shenoy grouped identified the ubiquitin conjugating enzyme UBE2L3 as a target of caspase-1. In this work, I have confirmed UBE2L3 as an indirect target of caspase-1 and characterised its role in inflammasomes-mediated immune responses. I show that UBE2L3 functions in the negative regulation of cellular pro-IL-1 via the ubiquitin- proteasome system. Following inflammatory stimuli, UBE2L3 assists in the ubiquitylation and degradation of newly produced pro-IL-1. However, in response to caspase-1 activation, UBE2L3 is itself targeted for degradation by the proteasome in a caspase-1-dependent manner, thereby liberating an additional pool of IL-1 which may be processed and released. UBE2L3 therefore acts a molecular rheostat, conferring caspase-1 an additional level of control over this potent cytokine, ensuring that it is efficiently secreted only in appropriate circumstances. These findings on UBE2L3 have implications for IL-1- driven pathology in hereditary fever syndromes, and autoinflammatory conditions associated with UBE2L3 polymorphisms. -

Celastrol Increases Glucocerebrosidase Activity in Gaucher Disease by Modulating Molecular Chaperones
Celastrol increases glucocerebrosidase activity in Gaucher disease by modulating molecular chaperones Chunzhang Yanga,1, Cody L. Swallowsa, Chao Zhanga, Jie Lua, Hongbin Xiaob, Roscoe O. Bradya,1, and Zhengping Zhuanga,1 aSurgical Neurology Branch, National Institute of Neurological Disorders and Stroke, National Institutes of Health, Bethesda, MD 20892-1260; and bInstitute of Chinese Materia Medica, China Academy of Chinese Medical Sciences, Beijing 100700, China Contributed by Roscoe O. Brady, November 19, 2013 (sent for review October 21, 2013) Gaucher disease is caused by mutations in the glucosidase, beta, acid increased the catalytic activity of mutant GCase. Celastrol interfered gene that encodes glucocerebrosidase (GCase). Glucosidase, beta, with the recruitment of Cdc37 to Hsp90 halting the assembly of the acid mutations often cause protein misfolding and quantitative loss requisite chaperone complex. Inhibition of Hsp90 reduced its rec- of GCase. In the present study, we found that celastrol, an herb de- ognition of mutant GCase and therefore limited the proteasomal rivative with known anticancer, anti-inflammatory, and antioxidant degradation of the mutant protein. Additionally, celastrol triggered activity, significantly increased the quantity and catalytic activity of a reorganization of the gene expression pattern of molecular chap- GCase. Celastrol interfered with the establishment of the heat-shock erones such as DnaJ homolog subfamily B members 1 and 9 protein 90/Hsp90 cochaperone Cdc37/Hsp90-Hsp70-organizing pro- (DNAJB1/9), heat shock 70kDa proteins 1A and 1B (HSPA1A/B), tein chaperone complex with mutant GCase and reduced heat-shock and Bcl2-associated athanogene 3 (BAG3). The presence of BAG protein 90-associated protein degradation. In addition, celastrol mod- family molecular chaperone regulator 3 (BAG3) further stabi- ulated the expression of molecular chaperones. -

Could Small Heat Shock Protein HSP27 Be a First-Line Target for Preventing Protein Aggregation in Parkinson’S Disease?
International Journal of Molecular Sciences Review Could Small Heat Shock Protein HSP27 Be a First-Line Target for Preventing Protein Aggregation in Parkinson’s Disease? Javier Navarro-Zaragoza 1,2 , Lorena Cuenca-Bermejo 2,3 , Pilar Almela 1,2,* , María-Luisa Laorden 1,2 and María-Trinidad Herrero 2,3,* 1 Department of Pharmacology, School of Medicine, University of Murcia, Campus Mare Nostrum, 30100 Murcia, Spain; [email protected] (J.N.-Z.); [email protected] (M.-L.L.) 2 Institute of Biomedical Research of Murcia (IMIB), Campus de Ciencias de la Salud, 30120 Murcia, Spain 3 Clinical & Experimental Neuroscience (NICE), Institute for Aging Research, School of Medicine, University of Murcia, Campus Mare Nostrum, 30100 Murcia, Spain; [email protected] * Correspondence: [email protected] (P.A.); [email protected] (M.-T.H.); Tel.: +34-868889358 (P.A.); +34-868883954 (M.-T.H.) Abstract: Small heat shock proteins (HSPs), such as HSP27, are ubiquitously expressed molecular chaperones and are essential for cellular homeostasis. The major functions of HSP27 include chaper- oning misfolded or unfolded polypeptides and protecting cells from toxic stress. Dysregulation of stress proteins is associated with many human diseases including neurodegenerative diseases, such as Parkinson’s disease (PD). PD is characterized by the presence of aggregates of α-synuclein in the central and peripheral nervous system, which induces the degeneration of dopaminergic neurons in the substantia nigra pars compacta (SNpc) and in the autonomic nervous system. Autonomic dys- function is an important non-motor phenotype of PD, which includes cardiovascular dysregulation, Citation: Navarro-Zaragoza, J.; among others. Nowadays, the therapies for PD focus on dopamine (DA) replacement. -
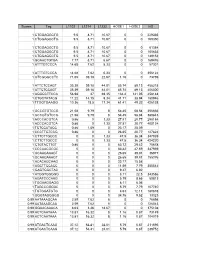
Scores Tag L1102 L1214 L1232 HOSE1 HOSE2 HS 1
Scores Tag L1102 L1214 L1232 HOSE1 HOSE2 HS 1 CTGGAGGCTG 9.5 8.71 10.67 0 0 229335 1 CTGGAGGCTG 9.5 8.71 10.67 0 0 169350 1 CTGGAGGCTG 9.5 8.71 10.67 0 0 61384 1 CTGGAGGCTG 9.5 8.71 10.67 0 0 105633 1 CTGGAGGCTG 9.5 8.71 10.67 0 0 149152 1 GCAACTGTGA 7.77 8.71 6.67 0 0 169476 1 ATTTGTCCCA 14.68 7.62 5.33 0 0 57301 1 ATTTGTCCCA 14.68 7.62 5.33 0 0 356122 1 GTCGGGCCTC 71.65 39.18 22.67 1.16 0 73769 1 ATTCTCCAGT 35.39 39.18 44.01 85.74 89.13 458218 1 ATTCTCCAGT 35.39 39.18 44.01 85.74 89.13 406300 1 AGGGCTTCCA 56.98 37 69.35 134.4 141.05 458148 1 CTGCTATACG 11.22 14.15 9.34 41.71 38.94 180946 1 TTGGTGAAGG 10.36 18.5 17.34 61.41 49.32 426138 1 GCCGTGTCCG 21.58 9.79 8 54.45 58.84 356666 1 GCCGTGTCCG 21.58 9.79 8 54.45 58.84 380843 1 ACCCACGTCA 0.86 0 1.33 27.81 20.77 298184 1 ACCCACGTCA 0.86 0 1.33 27.81 20.77 400124 1 TCTCCATACC 0.86 1.09 0 23.17 25.09 1 CCCTTGTCCG 0.86 0 0 26.65 20.77 127824 1 CTTCTTGCCC 0 0 1.33 47.5 36.34 347939 1 CTTCTTGCCC 0 0 1.33 47.5 36.34 424220 1 CTGTACTTGT 0.86 0 0 63.72 29.42 75678 1 CCCAACGCGC 0 0 0 83.42 47.59 347939 1 GCAAGAAAGT 0 0 0 26.65 39.81 36977 1 GCAAGAAAGT 0 0 0 26.65 39.81 155376 1 ACACAGCAAG 0 0 0 23.17 15.58 1 AGCTTCCACC 0 0 0 11.59 7.79 355542 1 GAGTGGCTAC 0 0 0 9.27 6.92 1 ATGGTGGGGG 0 0 0 8.11 22.5 343586 1 AGATCCCAAG 0 0 0 5.79 8.65 50813 1 TGGAAGGAGG 0 0 0 8.11 6.06 1 TAGCCGGGAC 0 0 0 5.79 7.79 107740 1 TGTGGATGTG 0 0 0 4.63 12.11 180878 1 GGGTAGGGGG 0 0 0 34.76 9.52 13323 0.99 AATAAAGCAA 2.59 7.62 8 0 0 76698 0.99 AATAAAGCAA 2.59 7.62 8 0 0 126043 0.99 GGAACAAACA 8.63 3.26 18.67 0 0 375108 -

(12) United States Patent (10) Patent No.: US 8.440,393 B2 Birrer Et Al
USOO8440393B2 (12) United States Patent (10) Patent No.: US 8.440,393 B2 Birrer et al. (45) Date of Patent: May 14, 2013 (54) PRO-ANGIOGENIC GENES IN OVARIAN OTHER PUBLICATIONS TUMORENDOTHELIAL CELL, SOLATES Boyd (The Basic Science of Oncology, 1992, McGraw-Hill, Inc., p. (75) Inventors: Michael J. Birrer, Mt. Airy, MD (US); 379). Tomas A. Bonome, Washington, DC Tockman et al. (Cancer Res., 1992, 52:2711s-2718s).* (US); Anil Sood, Pearland, TX (US); Pritzker (Clinical Chemistry, 2002, 48: 1147-1150).* Chunhua Lu, Missouri City, TX (US) Benedict et al. (J. Exp. Medicine, 2001, 193(1) 89-99).* Jiang et al. (J. Biol. Chem., 2003, 278(7) 4763-4769).* (73) Assignees: The United States of America as Matsushita et al. (FEBS Letters, 1999, vol. 443, pp. 348-352).* Represented by the Secretary of the Singh et al. (Glycobiology, 2001, vol. 11, pp. 587-592).* Department of Health and Human Abbosh et al. (Cancer Res. Jun. 1, 2006 66:5582-55.91 and Supple Services, Washington, DC (US); The mental Figs. S1-S7).* University of MD Anderson Cancer Zhai et al. (Chinese General Practice Aug. 2008, 11(8A): 1366 Center, Houston, TX (US) 1367).* Lu et al. (Cancer Res. Feb. 15, 2007, 64(4): 1757-1768).* (*) Notice: Subject to any disclaimer, the term of this Bagnato et al., “Activation of Mitogenic Signaling by Endothelin 1 in patent is extended or adjusted under 35 Ovarian Carcinoma Cells', Cancer Research, vol. 57, pp. 1306-1311, U.S.C. 154(b) by 194 days. 1997. Bouras et al., “Stanniocalcin 2 is an Estrogen-responsive Gene (21) Appl. -

Huang Lab Report
www.beatson.gla.ac.uk/danny_huang (Figure 2A). Upon DNA damage, Ser429 Figure 1 phosphorylation enhanced MDM2 Enzymatic cascade for Ub autoubiquitination and degradation in cells modifications explaining the rapid p53 activation. We also UBIQUITIN SIGNALLING described a strategy for preparation of MDM2 RING domain for structural analyses to enable rapid development MDM2 RING inhibitor. Insights into Deltex ubiquitin ligases Deltex (DTX) family E3s share a highly conserved C-terminal RING domain followed by a Deltex C-terminal domain (DTC). DTXs have been linked Post-translational modification with ubiquitin (Ub) initiated by to developmental processes involving Notch sequential actions of Ub-activating enzyme (E1), Ub-conjugating signalling and histone ubiquitination during DNA enzyme (E2) and Ub ligase (E3) regulates diverse cellular processes, damage repair. However, their functions remain largely unknown. We discovered that DTX E3s including signal transduction, cell cycle progression, apoptosis and harbour dual activities and catalyse both gene transcription. Deregulation in the Ub pathway is often ubiquitination of ADP-ribosylated substrate and ADP-ribosylation of Ub. We showed that DTX’s associated with human pathogenesis, including cancer. Our group C-terminal DTC domain harbour a conserved Group Leader uses structural biology and biochemical approaches to study the pocket that binds both ADP-ribose (ADPr) and NAD+ (Figure 2B). The DTC domains also bind Danny Huang enzymes in the Ub pathway to understand their regulation, levels of p53, the MDM2 mutant was able to poly-ADP-ribosylated substrates in cells. The restrain p53 activity sufficiently for normal Associate Scientist mechanistic functions and mutation-induced deregulation. We proximity of RING and DTC domain enables the growth. -

Airway Inflammation Via Ubch7 Ndfip1 Regulates Itch Ligase Activity
The Journal of Immunology Ndfip1 Regulates Itch Ligase Activity and Airway Inflammation via UbcH7 Mahesh Kathania,* Minghui Zeng,* Viveka Nand Yadav,† Seyed Javad Moghaddam,‡ Baoli Yang,x and K Venuprasad* The ubiquitin-ligating enzyme (E3) Itch plays a crucial role in the regulation of inflammation, and Itch deficiency leads to severe airway inflammation. However, the molecular mechanisms by which Itch function is regulated remain elusive. In this study, we found that nontypeable Haemophilus influenzae induces the association of Itch with Ndfip1. Both Itch2/2 and Ndfip12/2 mice exhibited severe airway inflammation in response to nontypeable Haemophilus influenza, which was associated with elevated expression of proinflammatory cytokines. Ndfip1 enhanced Itch ligase activity and facilitated Itch-mediated Tak1 ubiquitination. Mechanistically, Ndfip1 facilitated recruitment of ubiquitin-conjugating enzyme (E2) UbcH7 to Itch. The N-terminal region of Ndfip1 binds to UbcH7, whereas the PY motif binds to Itch. Hence, Ndfip1 acts as an adaptor for UbcH7 and Itch. Reconstitution of full-length Ndfip1 but not the mutants that fail to interact with either UbcH7 or Itch, restored the defect in Tak1 ubiquitination and inhibited elevated proinflammatory cytokine expression by Ndfip12/2 cells. These results provide new mechanistic insights into how Itch function is regulated during inflammatory signaling, which could be exploited therapeutically in inflammatory diseases. The Journal of Immunology, 2015, 194: 2160–2167. nflammation is essential for host defense against invading valent attachment of Ub to a conserved cysteine residue in the pathogens, such as viruses and bacteria (1). The inflammatory HECT domain, and this Ub is transferred to the target protein (5).