Notice of Intent to File a Lawsuit for Overdue Stock Assessment Reports
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

The Status of Wilderness Game in Alaska
PROCEEDDIGS OF THE FORTY-THIRD ANNUAL CONFERENCE OF WESTERN ASSOCIATION OF STA.TE GAME AND FISH COMMISSIONERS TUCSON, ARIZONA JUNE 10-12, 1963 -l~******** OFFICERS Robert J. Smith, President Ben Glading, Secretary Arizona California Walter J. Everin, 1st Vice President Montana Walter T. Shannon, 2nd Vice President California THE STATUS OF WILDERNESS GAME m .ALASKA ~ 1 ' JAMES W,. BROOKS .ALASKA DEP.AR'IMEJNT OF FISH .AND GAME The commonly held belief that wilderness types of wildlife cannot exist in close proximity to man or his land developments has helped to stim ulate the recent establishment of immense game refuges in Alaska and a contin uing agitation for additional wilderness areas and national parks. The belief stems from the early and permanent decl:lnes outside of Alaska suffered by wilderness species such as the timber wolf, grizzly bear, mountain sheep, and caribou which apparently coincided with the advance of civilization. In most cases it has been difficult or impossible to precisely identify proximate causes for such declines, though direct killing by man, diseases, change or usurption of habitat, or simply the presence of man or his cultural elements in the environment were undoubtedly involved. In .Alaska at this time we r...ave an opportunity to observe the reaction of various species and populations of wilderness types of wildlife to known influences. Our game research and man agement programs in most instances were initiated prior to significant hun1an intervention in the ecology of these forms •. The present status of wilderness types of :wildlife in Alaska often reflects the response of these animals to known levels and :types of exploi tation or other disrupting influences. -

WLD What Bear Goes Where MI Adaptation
DNRE - Wildlife Division Michigan Adaptation What Bear Goes Where? Teacher Plan Nuts and Bolts Objective: Students will [1] identify three species of bears and their habitats, and [2] generalize that animals have adapted in order to live where they do. Grade level: K – 4 Time: about 30 minutes Group size: Three groups of three to six students each. Setting: Indoors The major purpose of this activity is for students to recognize that animals can adapt to living in different environments, as shown in the example of three different kinds of bears. Materials Pictures of the three North American bear species, three large sheets of paper with the outline of one bear species on each and labeled, construction paper, pencils, scissors, glue. Background Information Polar bears have long necks, slender heads, and are white in appearance. They live along the Arctic coasts, mostly on the polar ice. The carnivorous polar bears feed mainly on fish and seals. Their thick fur keeps them warm, and the webbing between their toes makes them good swimmers. Unlike other bears, they have fur on the soles of their feet. Grizzly bears can dig up most of their food and can catch fish with their long claws. They also have a distinctive hump between their shoulders. They eat roots, tubers, gophers, marmots, and smaller rodents as well as carrion (rotten meat). They Adapted from Project Wild K-12 Curriculum and Activity Guide. Council for Environmental Education 2004. Lesson: What Bear Goes Where? DNRE - Wildlife Division Michigan Adaptation occasionally kill a larger animal for food. Grizzlies tend to live in the edges of forests but feed mostly in mountain meadows. -

© Clark Creative Education Wild Animal
© Clark Creative Education Wild Animal Conservation Relations, Patterns & Ideal Unit: Functions Time Range: 3-5 Days Supplies: Pencil & Paper Topics of Focus: - Creating Functions from Patterns - Rate of Change - Proportions Driving Question “How can conservation efforts save endangered species?” Culminating Experience A conservation plan Common Core Alignment: o Construct a function to model a linear relationship between two quantities. Determine the rate of change and initial value of the function from a description of a relationship or from two (x, y) 8.F.4 values, including reading these from a table or from a graph. Interpret the rate of change and initial value of a linear function in terms of the situation it models Calculate and interpret the average rate of change of a function (presented symbolically or as a F-IF.6 table) over a specified interval. Estimate the rate of change from a graph.★ F-BF.1 Write a function that describes a relationship between two quantities.★ F-LE.5 Interpret the parameters in a linear or exponential function in terms of a context. Interpret the slope (rate of change) and the intercept (constant term) of a linear model in the S-ID.7 context of the data. Procedures: A.) In “Animal Tracking”, students will practice proportions and use animal tracking mathematics to estimate the size of a population. B.) In “Endangered Species or Invasive Species?”, students will analyze patterns, write functions and predict the future. The population data is from the IUCN and WWF; however, numbers were slightly adjusted to make it an authentic pattern assignment. C.) In "Surviving Extinction”, students will use data to analyze pre-conservation and post-conservation data to answer questions about once critically endangered species. -

IUCN Briefing Paper
BRIEFING PAPER September 2016 Contact information updated April 2019 Informing decisions on trophy hunting A Briefing Paper regarding issues to be taken into account when considering restriction of imports of hunting trophies For more information: SUMMARY Dilys Roe Trophy hunting is currently the subject of intense debate, with moves IUCN CEESP/SSC Sustainable Use at various levels to end or restrict it, including through increased bans and Livelihoods or restrictions on carriage or import of trophies. This paper seeks to inform SpecialistGroup these discussions. [email protected] Patricia Cremona IUCN Global Species (such as large antlers), and overlaps with widely practiced hunting for meat. Programme It is clear that there have been, and continue to be, cases of poorly conducted [email protected] and poorly regulated hunting. While “Cecil the Lion” is perhaps the most highly publicised controversial case, there are examples of weak governance, corruption, lack of transparency, excessive quotas, illegal hunting, poor monitoring and other problems in a number of countries. This poor practice requires urgent action and reform. However, legal, well regulated trophy Habitat loss and degradation is a primary hunting programmes can, and do, play driver of declines in populations an important role in delivering benefits of terrestrial species. Demographic change for both wildlife conservation and for and corresponding demands for land for the livelihoods and wellbeing of indigenous development are increasing in biodiversity- and local communities living with wildlife. rich parts of the globe, exacerbating this pressure on wildlife and making the need for viable conservation incentives more urgent. © James Warwick RECOMMENDATIONS and the rights and livelihoods of indigenous and local communities, IUCN calls on relevant decision- makers at all levels to ensure that any decisions that could restrict or end trophy hunting programmes: i. -

Biological Opinion for the USFWS Region 7 Polar Bear and Pacific
Biological Opinion for the USFWS Region 7 Polar Bear and Pacific Walrus Deterrence Program Marine Mammals Management Office Anchorage, Alaska Prepared by: Fairbanks Fish and Wildlife Field Office U.S. Fish and Wildlife Service 101 12th Ave, Room 110 Fairbanks, AK 99701 January 13, 2014 1 Table of Contents List of Tables .................................................................................................................................. 3 Introduction ..................................................................................................................................... 3 The Proposed Action....................................................................................................................... 3 Requirements for Deterrence Authorization ............................................................................... 4 Application packet .................................................................................................................. 4 Availability of Biologist for Advice ........................................................................................... 6 Methods of Deterrence ................................................................................................................ 6 Polar Bears .............................................................................................................................. 6 Walruses .................................................................................................................................. 8 Who -

Powerpoint: Furbearer Regulations
2016-2017 and 2017-2018 Furbearer Trapping & Hunting Regulations Oregon Fish and Wildlife Commission June 9, 2016 Derek Broman 1 ODFW Carnivore-Furbearer Coordinator Presentation Overview License and Pelt Price Trends Species and Season Information Regulation Proposals • Sale of Unprotected Mammal Pelts • ODOT Crossing Structures 2 K.Kohl Furtaker License Sales Trends 1980-2015 6,000 Furtakers Furbearer Hunters 5,000 4,000 3,000 # Licenses 2,000 1,000 0 Year 3 New Furtaker License Trends 1996-2015 3,000 Returning Furtakers New Furtakers 2,500 2,000 1,500 # Licenses 1,000 500 0 Year 4 Furtaker Reporting Trend 2004-2014 100 Furtaker Furbearer Hunter 80 60 40 % Reporting % 20 0 Year 5 Average Pelt Prices 2014-2015 & 2015-2016 ─ Beaver ($14 $11) ─ Bobcat ($195 $211) ─ Coyote ($48 $25) AFWA 2015 Report ─ Marten ($28 $20) Average Trapping Expenses ─ Mink ($11 $6) US $1,694 ─ Muskrat ($5 $2) Oregon $1,761 ─ River Otter ($66 $60) ─ Raccoon ($6 $4) Based on Average Harvest ─ Gray Fox ($18 $11) and Prices for All Species: Western Oregon Furtaker: $1,564 ─ Red Fox ($26 $19) Eastern Oregon Furtaker: $2,006 Prices obtained from the Oregon 6 Territorial Council on Furs Species and Seasons 77 D. Budeau Season Recommendations Unprotected Mammals Open Entire Year for: Badger, Coyote, Nutria, Opossum, Porcupine, Skunks, Weasels Included in Harvest Report Protected Mammals No Open Season for: Fisher, Ringtail Cat, Wolverine, Kit Fox, Sea Otter 8 K.Kohl Furbearer Harvest Season Recommendations • No Changes Proposed to Season Dates for Trapping and Hunting Furbearers ─ Beaver Nov. 15 – Mar. 15 ─ Bobcat Dec. -

Marine Mammal Protection Act Complaint
1 Miyoko Sakashita (Cal. Bar # 239639) CENTER FOR BIOLOGICAL DIVERSITY 2 1212 Broadway, Suite 800 Oakland, CA 94612 3 Tel: (510) 844-7108 Fax: (510) 844-7150 4 Email: [email protected] 5 Lalli Venkatakrishnan (Cal. Bar # 323747) 6 CENTER FOR BIOLOGICAL DIVERSITY 1212 Broadway, Suite 800 7 Oakland, CA 94612 Phone: (510) 676-0348 8 Fax: (510) 844-7150 Email: [email protected] 9 10 Attorneys for Plaintiffs 11 UNITED STATES DISTRICT COURT NORTHERN DISTRICT OF CALIFORNIA 12 13 CENTER FOR BIOLOGICAL DIVERSITY, Civ. No. 14 a non-profit organization, and TURTLE ISLAND RESTORATION NETWORK, a COMPLAINT 15 non-profit organization; (Marine Mammal Protection Act, 16 Plaintiffs, 16 U.S.C. § 1361 et seq) 17 v. 18 SCOTT DE LA VEGA1 Acting Secretary, 19 United States Department of Interior; UNITED STATES FISH AND WILDLIFE 20 SERVICE; 21 Defendants. 22 23 24 25 26 27 28 1 Pursuant to Fed. R. Civ. P. 25(d), David Bernhardt’s successor is automatically substituted as a party when Mr. Bernhardt ceases to hold office as Secretary of the U.S. Department of Interior, and any misnomer not affecting the parties’ substantial rights must be disregarded. 1 INTRODUCTION 2 1. This is an action challenging the failure of Defendants Scott de la Vega, Acting 3 Secretary of the Interior, and the United States Fish and Wildlife Service (collectively “the 4 Service”) to comply with their non-discretionary obligations under the Marine Mammal 5 Protection Act (“MMPA”). 16 U.S.C. § 1361, et seq. Specifically, the Service has failed to issue 6 updated stock assessment reports for marine mammals under its jurisdiction—sea otters, polar 7 bears, walruses, and manatees—within the timeframes mandated by the statute. -

Modern Status of Sea Otter Population on the Commander Islands
Modern status of Sea otter population on the Commander Islands Alexander Burdin, Kamchatka branch of Pacific Institute of Geography, RAS, University of Alaska, Fairbanks, ASLC, Sergey Zagrebelny, Commander preserve Commander Population declines Islands (population growing) Background Only 200 nm strait between Commander Island and western Aleutian islands. Dramatic decline of sea otter populations (up to 90% and more reduction), and some other marine mammal species (harbor seal, SSL) across the Aleutian Archipelago and Alaska Peninsula during past several decades. Increasing (13%/year) sea otter population on the Commander Islands. Research Objectives Commander-Aleutian islands c comparisonsomparisons ¾ to better understand ultimate reasons for the decline ¾ to characterize physiology, behavior, and demography of sea otter population near K. ¾ to expand studies of sea otter-kelp forest interactions 2004-2005 winter field work Sea otter mortality, disease, physical conditions monitoring: Carcasses collection / capture : Age/sex composition Necropsy: disease, causes of death, virology, female reproductive tracts, parasites, stomach contents Biosampling 2006 summer work on the Bering Island Totally 33 sea otters were caught using nets and release. 2006 summer work on the Bering Island 27 sea otters (22 females and 5 males) were instrumented with TDR and radio tag Ongoing investigation and future research Goal 1. Ecosystem research: long-term changes in near shore communities under sea otter predation. availability and abundance of sea otter food recourses. analysis of TDR’s data. direct observation on feeding sea otters. scat analysis. Retrospective analysis of sea otter feeding habits on the Commanders. Ongoing investigation and future research Goal II. Monitoring of sea otter population: annual survey (skiff and shore based). -

Polar Bear (Ursus Maritimus) Care Manual
Polar Bear (Ursus maritimus) Care Manual POLAR BEAR (Ursus maritimus) CARE MANUAL CREATED BY THE AZA Polar Bear Species Survival Plan® IN ASSOCIATION WITH THE AZA Bear Taxonomic Advisory Group Association of Zoos and Aquariums 1 Polar Bear (Ursus maritimus) Care Manual Polar Bear (Ursus maritimus) Care Manual Published by the Association of Zoos and Aquariums in association with the AZA Animal Welfare Committee Formal Citation: AZA Bear TAG 2009. Polar Bear (Ursus maritimus) Care Manual. Association of Zoos and Aquariums, Silver Spring, MD. Original Completion Date: 16 December 2009 Authors and Significant Contributors: See Appendix J AZA Staff Editors: Dr. Debborah Colbert, AZA Vice President, Conservation & Science Reviewers: Dr. Randi Meyerson (ACM Coordinator, Toledo Zoo) Dr. Joseph C.E. Barber, (AZA, ACM Consultant) Dr. Don Moore (SI-NZP, AZA Bear TAG Advisor) Dr. Barbara Kohn (APHIS) Dr. Steven Amstrup (USGS) Dr. Lydia Kolter (EAZA Bear TAG Chair) Barbara Henry, M.S. (AZA Nutrition SAG, Curator of Nutrition, Cincinnati Zoo & Botanical Garden) Ann Ward, M.S. (AZA Nutrition SAG, Director, Nutritional Services for Fort Worth Zoo) Cover Photo Credit: Randi Myerson Disclaimer: This manual presents a compilation of knowledge provided by recognized animal experts based on the current science, practice, and technology of animal management. The manual assembles basic requirements, best practices, and animal care recommendations to maximize capacity for excellence in animal care and welfare. The manual should be considered a work in progress, since practices continue to evolve through advances in scientific knowledge. The use of information within this manual should be in accordance with all local, state, and federal laws and regulations concerning the care of animals. -

Natural History of the Southern Sea Otter
Natural History of the Southern Sea Otter C Compiled by Gena Bentall 2017 Description Sea otters are members of the weasel or mustelid family. Like other members of this family, they have very thick fur. In fact, at 850,000 to one million hairs per square inch, they have the thickest fur of any mammal. Their fur consists of two types of hairs, interlocking underfur (which provides insulation) and longer guard hairs (that help water run off the coat). This system traps a layer of air next to their skin so, when fur is well groomed, their skin does not come in contact with sea water. Sea otters are usually dark brown, and some individuals may be progressively lighter colored (grizzled) on the head, neck, chest and forearms due to loss of pigmentation in the guard hairs. Extent of grizzle can be related to age and individual variation. Sea otters are the smallest marine mammal, and with their flipper-shaped hind feet are well adapted to a marine environment. In California adult females weigh 35-60 pounds (16-27 kg); males reach up to 90 pounds (40 kg). Alaskan sea otters are bigger with males weighing as much as 100 pounds (45 kg). Range/Habitat Sea otters once ranged around the North Pacific Rim from Mexico through Alaska, Russia, and Japan. The maritime fur trade of the 1700-1800s brought sea otters to the brink of extinction and fragmented the once continuous population. There are currently 3 subspecies of sea otter, the Northern Sea Otter (Enhydra lutris kenyoni), the Asian, or Russian, Sea Otter (Enhydra lutris lutris) and our Southern, or California, Sea Otter (Enhydra lutris nereis). -
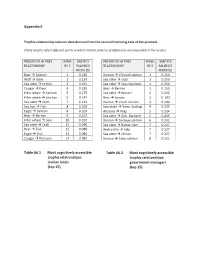
Combining Salience and Network Analyses To
Appendix 6 Trophic-relationship salience data derived from the second freelisting task of the protocol (Table lengths reflect different points at which notable patterns of differences are observable in the results) PREDATOR Æ PREY RANK SMITH’S PREDATOR Æ PREY RANK SMITH’S RELATIONSHIP BY S SALIENCE RELATIONSHIP BY S SALIENCE INDEX (S) INDEX (S) Bear Æ Salmon 1 0.230 Human Æ Chinook salmon 1 0.250 Wolf Æ Deer 2 0.219 Sea otter Æ crab 1 0.250 Sea otter Æ Urchin 3 0.191 Sea otter Æ Sea cucumber 1 0.250 Cougar Æ Deer 4 0.185 Bear Æ Berries 1 0.250 Killer whale Æ Salmon 5 0.175 Sea otter Æ Mussel 2 0.242 Killer whale Æ Sea lion 6 0.147 Bear Æ Leaves 2 0. 242 Sea otter Æ Clam 7 0.133 Human Æ Chum salmon 3 0.240 Sea lion Æ Fish 8 0.109 Sea otter Æ Swm. Scallop 4 0.239 Eagle Æ Salmon 8 0.109 Abalone Æ Kelp 5 0.234 Bear Æ Berries 9 0.107 Sea otter Æ Gnk. Barnacle 5 0.234 Killer whale Æ Seal 10 0.103 Human Æ Sockeye salmon 6 0.231 Sea otter Æ Crab 11 0.090 Sea otter Æ Butter clam 7 0.227 Bear Æ Fish 12 0.088 Red urchin Æ Kelp 7 0.227 Eagle Æ Fish 13 0.086 Sea otter Æ Chiton 7 0.227 Cougar Æ Raccoon 14 0.082 Human Æ Coho salmon 8 0.221 Table A6.1 Most cognitively accessible Table A6.2 Most cognitively accessible trophic relationships: trophic relationships: civilian locals government managers (top 15) (top 15) PREDATOR Æ PREY RANK SMITH’S PREDATOR Æ PREY RANK SMITH’S RELATIONSHIP BY S SALIENCE RELATIONSHIP BY S SALIENCE INDEX (S) INDEX (S) Sea otter Æ Urchin 1 0.264 Killer whale Æ Salmon 1 0.270 Sea otter Æ Clam 2 0.224 Bear Æ Salmon 2 0.267 Wolf Æ Deer 3 0.198 Wolf Æ Deer 3 0.236 Bear Æ Salmon 4 0.184 Cougar Æ Deer 4 0.189 Cougar Æ Deer 5 0.180 Killer whale Æ Sea lion 5 0.138 Bear Æ Berries 6 0.163 Sea otter Æ Urchin 6 0.134 Killer whale Æ S. -

PUP 681: a SEA OTTER RESCUE STORY Curriculum and Storytime Guide
Pup 681: Storytime andA StoryCurriculumtime Guideand Curriculum Guide for 1 Pup 681: A Sea Otter Rescue Story by Jean Reidy, illustrated by Ashley Crowley PUP 681: A SEA OTTER RESCUE STORY ©2019 by Jean Reidy; Art by Ashely Crowley; Henry Holt and Company Praise for Pup 681 “Unabashedly adorable” ~ Kirkus Reviews, starred review “...otter enthusiasts will likely enjoy the tender relationship between 681 and her attentive caregiver.” ~ Publishers Weekly Learning activities align with CCSS (Common Core State Standards) and NGSS (Next Generation Science Standards) where applicable. Guide created by Natalie Lorenzi (www.nataliediaslorenzi.com) Artwork ©Ashely Crowley Pup 681: Storytime and Curriculum Guide 2 Story Summary Washed ashore alone, a tiny sea otter pup needs help! Soon, a rescuer is there to take her in and keep her warm and fed. The pup faces challenges in her new life without her sea otter family. But with the love and care of her rescuer, she flourishes in her new home. Inspired by a true story, this is a heartwarming and hopeful tale about family and love. About the Author Jean Reidy’s fun, lively and award-winning picture books have earned their spots as favorites among readers and listeners of all ages and from all over the world. She is a frequent presenter on writing and reading and at schools and storytimes across the country—in person and virtually. Jean is a member of the Society of Children’s Book Writers and Illustrators, the National Council of Teachers of English, the Colorado Council International Reading Association and she has served on the board of Reach Out and Read Colorado.