<I>Coursetia</I> (Leguminosae) from Eastern Brazil
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
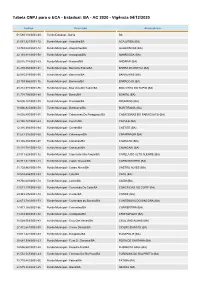
Tabela CNPJ Para O ECA - Estadual: BA - AC 2020 - Vigência 04/12/2020
Tabela CNPJ para o ECA - Estadual: BA - AC 2020 - Vigência 04/12/2020 Código Descrição Abrangência 01.580.116/0001-00 Fundo Estadual - Bahia BA 21.551.921/0001-12 Fundo Municipal - Acajutiba/BA ACAJUTIBA (BA) 14.784.004/0001-12 Fundo Municipal - Alagoinhas/BA ALAGOINHAS (BA) 22.103.811/0001-50 Fundo Municipal - Amargosa/BA AMARGOSA (BA) 20.615.114/0001-53 Fundo Municipal - Andaraí/BA ANDARAI (BA) 21.270.868/0001-81 Fundo Municipal - Barra Da Estiva/BA BARRA DA ESTIVA (BA) 22.915.018/0001-56 Fundo Municipal - Barreiras/BA BARREIRAS (BA) 29.103.966/0001-36 Fundo Municipal - Barrocas/BA BARROCAS (BA) 28.312.971/0001-96 Fundo Municipal - Boa Vista Do Tupim/BA BOA VISTA DO TUPIM (BA) 31.738.706/0001-89 Fundo Municipal - Bonito/BA BONITO (BA) 16.595.151/0001-70 Fundo Municipal - Brumado/BA BRUMADO (BA) 18.006.845/0001-50 Fundo Municipal - Buritirama/BA BURITIRAMA (BA) 18.692.450/0001-58 Fundo Municipal - Cabaceiras Do Paraguaçu/BA CABACEIRAS DO PARAGUACU (BA) 23.140.101/0001-63 Fundo Municipal - Caculé/BA CACULE (BA) 12.316.993/0001-94 Fundo Municipal - Caetité/BA CAETITE (BA) 31.677.315/0001-00 Fundo Municipal - Cafarnaum/BA CAFARNAUM (BA) 63.186.456/0001-08 Fundo Municipal - Camacan/BA CAMACAN (BA) 18.118.784/0001-12 Fundo Municipal - Camaçari/BA CAMACARI (BA) 23.811.628/0001-72 Fundo Municipal - Capela do Alto Alegre/BA CAPELA DO ALTO ALEGRE (BA) 28.941.344/0001-14 Fundo Municipal - Capim Grosso/BA CAPIM GROSSO (BA) 31.720.861/0001-78 Fundo Municipal - Castro Alves/BA CASTRO ALVES (BA) 12.574.686/0001-03 Fundo Municipal - Catu/BA CATU (BA) -

ISSN 1677-7069 3 Nº 177, Segunda-Feira, 15 De Setembro De 2014
174 ISSN 1677-7069 3 Nº 177, segunda-feira, 15 de setembro de 2014 ESTADO DO AMAZONAS PREFEITURA MUNICIPAL DE PARINTINS AVISOS DE LICITAÇÃO PREFEITURA MUNICIPAL DE APUÍ CONCORRÊNCIA No- 7/2014 AVISO DE HOMOLOGAÇÃO E ADJUDICAÇÃO EXTRATO TERMO ADITIVO PREGÃO PRESENCIAL Nº 8/2014-CML Objeto: CONTRATAÇÃO DE EMPRESA ESPECIALIZADA PARA A PRESTAÇÃO DE SERVIÇOS DE MANUTENÇÃO PREDIAL SEGUNDO TERMO ADITIVO DO TERMO DE CONTRATO Nº. CONSIDERANDO o relatório apresentado pela Pregoeira e DAS UNIDADES ESCOLARES DA REDE MUNICIPAL DE EN- 002/2013 OBJETO: AQUISIÇÃO DE COMBUSTÍVEL E LUBRI- equipe de apoio no Processo nº 045/2014, relativo à Licitação do tipo SINO DE ALAGOINHAS - BAHIA. Tipo: Menor preço global. Re- FICANTES para recuperação de estradas e vicinais conforme Termo Pregão Presencial do Registro de Preços nº 8/2014, CONSIDERAN- cebimento de envelopes e abertura no dia 21/10/2014, às 08h30min. de Convênio nº. 03/2013 - IDAM), objetivando a ampliação da in- DO ainda, a inexistência de qualquer recurso pendente ao referido O Edital estará á disposição na COPEL situada no CAM - CENTRO ADMINISTRATIVO MUNICIPAL, Rua Manoel Romão, N° 23, Ala- fraestrutura produtiva da Zona Rural do Município de Apuí. CE- procedimento licitatório e o que mais consta dos autos do men- cionado processo, HOMOLOGO a deliberação da Pregoeira e Equipe goinhas Velha, Alagoinhas/BA ao preço de R$ 100,00 (cem reais). LEBRANTES: Prefeitura Municipal de Apuí, como Município Con- Dados Bancários: Banco do Brasil - AG: 0158-9, C/C:16.658-8.. tratante, e a empresa. MAFUI AGRO INDUSTIAL E COMERCIAL, de Apoio, constante no relatório supracitado para todos os efeitos Maiores informações pelo Tel. -

Revista Vc 7 N 1.Pmd
Marco Antônio Sartori, Fátima Machado de Souza Lima, RonaldoISSN Perez, 1679-1614 Mauro Nacif Rocha & Danielle Dias Sant’Anna Martins PROPOSTA LOCACIONAL PARA UNIDADES DE PROCESSAMENTO DE MAMONA PELA AGRICULTURA FAMILIAR, NO CENTRO NORTE DO ESTADO DA BAHIA1 Marco Antônio Sartori2 Fátima Machado de Souza Lima3 Ronaldo Perez4 Mauro Nacif Rocha5 Danielle Dias Sant’Anna Martins6 Resumo: Este trabalho avalia locais para instalação de unidades de produção de óleo vegetal de mamona na região centro-norte do estado da Bahia. Na região existem 55 municípios produtores de mamona, e a cidade de Candeias será o destino do óleo vegetal produzido, em razão da instalação da unidade de produção de biodiesel da PETROBRAS. Com a finalidade de garantir a participação da agricultura familiar, o estudo considera unidades de processamento de pequena escala, com capacidades de processamento de 7,5 e 22,5 toneladas/dia de mamona. Para a solução, desenvolveu-se um modelo matemático, utilizando o software XPRESS MP Release, 2007, considerando as produções nos municípios, as distâncias entre as áreas de produção agrícola e as cidades candidatas ao esmagamento, e destas até a usina de produção de biodiesel. Como resultado, obteve-se a indicação de oito cidades potenciais para instalação de plantas de produção de óleo vegetal, de forma a minimizar o custo logístico. Palavras-chave: Localização, óleo de mamona, custos de transporte. 1 Recebido em 02/09/08; Aceito em 28/11/08. 2 Doutorando em Ciência e Tecnologia de Alimentos/UFV. E-mail: [email protected] 3 Mestranda em Ciência e Tecnologia de Alimentos/UFV. E-mail: [email protected] 4 Professor do Departamento de Tecnologia de Alimentos da Universidade Federal de Viçosa. -

Boletim Epidemiológico Bahia – COVID-19
Nº 399 - 27/04/2021 Situação Epidemiológica Atual Na Bahia, nas últimas 24 horas, foram registrados 4.076 casos de Covid-19 (taxa de crescimento de +0,5%) e 3.494 recuperados (+0,4%). Dos 889.931 casos confirmados desde o início da pandemia, 856.110 já são considerados recuperados, 15.627 encontram- se ativos e 18.194 tiveram óbito confirmado. Foram considerados para os casos confirmados: critérios laboratoriais (RT-PCR, imunológicos e teste rápidos), critérios clínico-epidemiológico e clínico imagem. Tabela 1. Distribuição dos casos de COVID-19, segundo situação da investigação. Bahia, 2020-2021*. Classificação Casos N % Confirmados laboratorialmente RT-PCR 531458 23,35 Confirmados laboratorialmente imunológico 3073 0,14 Confirmados laboratorialmente teste rápido 346137 15,21 Confirmados clínico epidemiológico 6052 0,27 Confirmados clínico imagem 1120 0,05 Aguardando validação dos municípios* 2091 0,09 Total 889931 39,11 Descartados 1191910 52,38 Em investigação 193784 8,52 Total 2275625 100 *Casos confirmados de covid-19 cuja condição clinica permanece sendo acompanhada ou aguardando autorização pelos municipios. Fonte: e-SUS-Ve/ Divep e GAL/Lacen, dados obtidos em 27/04/2021. Figura 1. Distribuição do acumulado de casos confirmados por Covid-19 por semana epidemiológica Bahia, 2020-2021.** Na Figura 1, observa-se que da 14ª Semana Epidemiológica (SE) de 2020 a 16º SE de 2021 houve um aumento do número de casos confirmados de COVID-19 na Bahia, com incremento de 262.043,75% de casos nesse período. Página 2/19 Até 27/04/2021 no Estado da Bahia, o coeficiente de incidência foi de 5.983,51/100.000 habitantes. -

Boletim Epidemiológico Bahia – COVID-19
Nº 97 - 29/06/2020 Situação Epidemiológica Atual Na Bahia, nas últimas 24 horas, foram registrados 972 casos de Covid-19 (taxa de crescimento de +1,4%), 52 óbitos (+3,0%) e 874 curados (+2,0%). Dos 69.467 casos confirmados desde o início da pandemia, 43.946 já são considerados curados, 23.721 encontram-se ativos e 1.800 tiveram óbito confirmado. Para todos os casos confirmados foram considerados os resultados laboratoriais, teste rápido e clínico-epidemiológico. Do total de casos confirmados para o Estado, 46.369 (15,42%) foram confirmados laboratorialmente, 1.810 (0,60%) pelo critério clínico-epidemiológico, 20.827 (6,93%) por teste rápido e 461 (0,15 %) confirmados aguardando validação do município. O total de casos notificados foram 300.738, desses 69.467 (23,10%) são casos confirmados, 159.204 (52,94%) são descartados e 72.067 (23,96%) permanecem em investigação epidemiológica (Tabela 1). Tabela 1. Distribuição dos casos de COVID-19, segundo situação da investigação. Bahia, 2020*. Classificação Casos N % Confirmados laboratorialmente 46369 15,42 Confirmados clínico epidemiológica 1810 0,60 Confirmados teste rápido 20827 6,93 Aguardando validação dos municípios* 461 0,15 Total 69467 23,10 Descartados 159204 52,94 Em investigação 72067 23,96 Total 300738 100 *Casos confirmados de covid-19 cuja condição clinica permanece sendo acompanhada ou aguarda autorização pelos municipios. Fonte: e-SUS-Ve/ Divep e GAL/Lacen, dados obtidos em 22/06/2020, sujeito a alterações. A plataforma REDCAP não permitiu mais a inclusão de novos casos suspeitos da COVID-19 desde o dia 27/03/2020, assim as notificações de novos casos passaram a ser feitas na nova plataforma (http://notifica.saude.gov.br). -

Subchefia Para Assuntos Jurídicos
Presidência da República Casa Civil Subchefia para Assuntos Jurídicos DECRETO DE 11 DE JUNHO DE 2010. Cria o Parque Nacional e o Refúgio de Vida Silvestre de Boa Nova, no Estado da Bahia, e dá outras providências. O PRESIDENTE DA REPÚBLICA, no uso da atribuição que lhe confere o art. 84, inciso IV, da Constituição, e tendo em vista o disposto nos arts. 11 e 13 da Lei no 9.985, de 18 de julho de 2000, e o que consta do Processo no 02001.006918/2005-09, DECRETA: Art. 1o Ficam criados o Parque Nacional de Boa Nova, com aproximadamente 12.065 ha, e o Refúgio de Vida Silvestre de Boa Nova, com aproximadamente 15.024 ha, localizados nos Municípios de Boa Nova, Manoel Vitorino e Dário Meira, no Estado da Bahia, cujos objetivos são: I - proteger integralmente e regenerar os ecossistemas naturais da transição entre Mata Atlântica e Caatinga, especialmente a Mata-de-Cipó; II - garantir a manutenção de populações viáveis de espécies de aves e mamíferos ameaçadas de extinção, especialmente o gravatazeiro (Rhopornis ardesiacus); III - manter e recuperar mananciais e cursos d'água; IV - possibilitar o desenvolvimento de atividades de educação e interpretação ambiental, recreação em contato com a natureza e turismo ecológico; e V - possibilitar o desenvolvimento de pesquisa científica. Parágrafo único. O Refúgio de Vida Silvestre de Boa Nova tem também por objetivo proporcionar conectividade entre as áreas do Parque Nacional de Boa Nova. Art. 2o O Parque Nacional de Boa Nova tem seus limites descritos a partir da base cartográfica elaborada pela Superintendência do Desenvolvimento do Nordeste - SUDENE, Cartas Manoel Vitorino (SD- 24-Y-B-I) e Poções (SD-24-Y-B-IV), na escala 1:100.000, publicadas em Projeção Universal Transversa de Mercator, Fuso 24, Datum Horizontal Córrego Alegre, com os seguintes memoriais descritivos: I - Área 1: inicia-se a descrição do perímetro da Área 1 a partir do ponto 1, localizado no entroncamento de estradas sem denominação, uma delas derivando da BR-030, de c.p.a. -

BOLETIM SEGOV MUNICIPAL Articulação Federativa Para O Enfrentamento Da Situação De Emergência Decorrente Do Coronavírus (Covid-19)
417 municípios 4ª Edição Bahia BOLETIM SEGOV MUNICIPAL Articulação Federativa para o enfrentamento da situação de emergência decorrente do Coronavírus (Covid-19) Presidência da República - Secretaria de Governo Secretaria Especial de Assuntos Federativos 15 de julho de 2020 BAHIA - Entregas do Governo Federal Ações e Serviços Públicos de Saúde - Insumos Estratégicos Equipamentos de Testes Medicamentos Proteção Individual (EPI’s) Máscara Cirúrgica Máscara N95 RT-PCR Cloroquina 3.741.400 218.500 137.392 81.000 Álcool em gel (L) Óculos e protetor 33.499 facial 101.840 Rápidos Oseltamivir Sapatilha e 509.020 214.170 Avental Luvas Touca 198.299 1.666.496 1.102.800 */ Consulta em 13 de julho de 2020 no portal do Ministério da Saúde: https://covid-insumos.saude.gov.br/paineis/insumos/painel_leitos.php Elaboração: SEAF/SEGOV/PRESIDÊNCIA DA REPÚBLICA 2 BAHIA - Entregas do Governo Federal Ações e Serviços Públicos de Saúde - Insumos Estratégicos Respiradores Leitos Distribuídos Leitos Locados Leitos UTI Habilitados 40 539 321 Leitos UTI Adulto Leitos UTI Adulto SUS não SUS 869 704 */ Consulta em 13 de julho de 2020 no portal do Ministério da Saúde: https://covid-insumos.saude.gov.br/paineis/insumos/painel_leitos.php Elaboração: SEAF/SEGOV/PRESIDÊNCIA DA REPÚBLICA 3 BAHIA - Repasses de Recursos do Governo Federal FNS - Fundo Nacional Apoio FPE Apoio FPM PFEC - Programa de Saúde (FAF-COVID) (MP 938) (MP 938) Federativo (Março, Abril, Maio, Junho e (Abril, Maio, Junho e Julho) (Abril, Maio, Junho e Julho) (LC n° 173) Julho) (Junho e Julho) R$ 381,80 -

Seção 3 ISSN 1677-7069 Nº 229, Quarta-Feira, 27 De Novembro De 2019
Seção 3 ISSN 1677-7069 Nº 229, quarta-feira, 27 de novembro de 2019 PREFEITURA MUNICIPAL DE CARAVELAS PREFEITURA MUNICIPAL DE IBOTIRAMA EXTRATO DE CONTRATO Nº CP 005-2019 AVISO DE LICITAÇÃO TOMADA DE PREÇOS Nº 4/2019-CPL/PMI/BA Extrato de Contrato Nº CP 005-2019. Processo de Licitação da Chamada Pública nº 005- 2019. OBJETO: Fornecimento parcelado de Gêneros Alimentícios da Agricultura Familiar e A Comissão de Licitação da Prefeitura Municipal de Ibotirama comunica aos do Empreendedor Familiar Rural. CONTRATADA: ASSOCIAÇÃO DOS PRODUTORES RURAIS DA NOVA CRAVILINA, com o valor do projeto R$ 6.728,75 (seis mil, setecentos e vinte e interessados que se encontra aberta a licitação na modalidade TOMADA DE PREÇOS Nº oito reais, setenta e cinco centavos). Vigência: até 30/12/2019. Caravelas/Ba, 26/11/19 - 004/2019-CPL/PMI/BA, do tipo empreitada por menor preço global, tendo como objeto a Prefeito Municipal. contratação de empresa especializada em obras e serviços de engenharia para construção PREFEITURA MUNICIPAL DE CASTRO ALVES da cobertura da praça da feira municipal de Ibotirama-BA. Será realizada no dia 13/12/2019, às 08:30 h, na sala de Licitações desta Prefeitura. O edital e as informações AVISOS DE LICITAÇÃO serão obtidos na sede da Prefeitura situada na Praça Ives de Oliveira, 78 - Centro - PREGÃO PRESENCIAL Nº 40/2019 Ibotirama - BA, das 8:00 às 12:00 h ou pelo telefone (77) 3698-1512. A Pregoeira realizará reabertura do PP n° 040/2019 PA nº 071/2019, Tipo: NEYLON NICHELLE PEREIRA DE SÁ TELES Menor PREÇO POR ITEM. -

ISSN 1677-7069 3 Nº 66, Segunda-Feira, 8 De Abril De 2013
196 ISSN 1677-7069 3 Nº 66, segunda-feira, 8 de abril de 2013 PREFEITURA MUNICIPAL DE ARACATU PREFEITURA MUNICIPAL DE IPIRÁ PREGÃO PRESENCIAL No- 31/2013 AVISO DE REVOGAÇÃO AVISOS LICITAÇÃO o O Município de Jequié torna público que no dia 18/04/2013, o PREGÃO PRESENCIAL N- 22/2013 PREGÃO PRESENCIAL N- 11 / 2 0 1 3 às 14:00 horas, na Praça Duque de Caxias, s/n, no Prédio da Pre- feitura Municipal, estará realizando sessão de PREGÃO PRESEN- Comunica que por motivo de modificações no Edital a pe- CIAL destinado à AQUISIÇÃO DE MEDICAMENTOS PARA Objeto: Contratação de empresa especializada para prestar Serviços dido do Ministério Público através da Promotora de Justiça Drª Clara ATENDER AS NECESSIDADES DA SECRETARIA DE SAÚDE de Gestão dos Serviços de Saúde Hospitalar, Atenção Básica, Vi- Akemi Kobayashi Koontz, revoga o Pregão Presencial 22/2013, para DO MUNICÍPIO CONFORME AS ESPECIFICAÇÕES NO ANEXO gilância à Saúde, e SAMU no Município, sob o regime de menor contratação de empresa especializada em locação de veículos des- I DO EDITAL 31/2013. O edital e seus elementos poderão ser ad- preço global. Abertura: 18/04/13, às 08:00 horas. Edital: Sede da tinados ao transporte escolar, que seria realizado no dia 08/04/2013 às quiridos na Praça Duque de Caxias, s/n no Prédio da Prefeitura. Prefeitura Municipal, situada à Rua Libério Maia, 37 - Aracatu - BA, 10 hs. PREGÃO PRESENCIAL No- 32/2013 46130-000, das 08:30 às 11:30 hs. Ipirá-BA, 5 de abril de 2013. o MURILO TADEU DA SILVA LIMA O Município de Jequié torna público que no dia 19/04/2013, PREGÃO -

Of the State of Bahia, Brazil 99 Doi: 10.3897/Zookeys.517.10219 RESEARCH ARTICLE Launched to Accelerate Biodiversity Research
A peer-reviewed open-access journal ZooKeys 517:New 99–110 species (2015) and new records in Cerambycidae (Coleoptera) of the state of Bahia, Brazil 99 doi: 10.3897/zookeys.517.10219 RESEARCH ARTICLE http://zookeys.pensoft.net Launched to accelerate biodiversity research New species and new records in Cerambycidae (Coleoptera) of the state of Bahia, Brazil Maria Helena M. Galileo1,3, Ubirajara R. Martins†, Antonio Santos-Silva2 1 PPG Biologia Animal, Departamento de Zoologia, Universidade Federal do Rio Grande do Sul, Porto Alegre, RS, Brazil 2 Museu de Zoologia, Universidade de São Paulo, Caixa Postal 42594, 04218-970, São Paulo, São Paulo, Brazil 3 Fellow of the Conselho Nacional de Desenvolvimento Científico e Tecnológico Corresponding authors: Maria Helena M. Galileo ([email protected]); Antonio Santos-Silva ([email protected]) Academic editor: S. Lingafelter | Received 28 June 2015 | Accepted 3 August 2015 | Published 12 August 2015 http://zoobank.org/7AD74C1C-B9A7-4EAC-852E-BA1571A6884F Citation: Galileo MHM, Martins UR, Santos-Silva A (2015) New species and new records in Cerambycidae (Coleoptera) of the state of Bahia, Brazil. ZooKeys 517: 99–110. doi: 10.3897/zookeys.517.10219 Abstract Two new species are described from Bahia (Brazil): Coleoxestia beckeri (Cerambycini), and Oncioderes obliqua (Onciderini). Nine species are recorded for the first time for Bahia (Brazil). Key to species of Oncioderes Martins & Galileo, 1990 is provided. Coleoxestia beckeri is included in a previous key. Keywords Neotropical, new records, new species, taxonomy Introduction The study of the specimens sent by Vitor O. Becker from the state of Bahia in Brazil allowed the description of two new species, and also new records for the state. -

Matrícula Da Rede Estadual De Ensino 22 De Março a 14 De Abril De 2021
Matrícula da rede estadual de ensino 22 de março a 14 de abril de 2021 BLOCO 01 (22 a 25/03/2021) NTE: 01, 02, 03, 04, 05, 06 e 08 1.Transferência de Estudantes da Rede Estadual: ▪ Para os estudantes matriculados no ano letivo de 2020, ao qual se aplique uma das seguintes situações: - A escola não oferece o ano/série subsequente; - Mudança de domicílio. Data: 22 a 25/03 ▪ Municípios: América Dourada - Abaíra - Almadina - Andaraí - Arací- Arataca - Aratuípe - Aurelino Leal - Barra - Barra da Estiva - Barra do Mendes - Barro Alto - Barro Preto - Barrocas - Biritinga - Bom Jesus da Lapa - Boninal - Bonito - Brotas de Macaúbas - Buerarema - Caatiba - Cafarnaum - Cairu - Camacan - Camamu - Canarana - Canavieiras - Candeal - Cansanção - Carinhanha - Central - Coaraci - Conceição do Coite - Feira da Mata - Firmino Alves - Floresta Azul - Gandu - Gentio do Ouro - Ibicaraí - Ibicoara - Ibicuí - Ibipeba - Ibirapitanga - Ibitiara - Ibitita - Ibotirama - Ichu - Igapora - Igrapiuna - Iguaí - Ilheus - Ipupiara - Iramaia - Iraquara - Irecê - Itabuna - Itacaré - Itaetê - Itaguaçu da Bahia - Itaju do Colônia - Itajuipe - Itambé - Itapé - Itapetinga - Itapitanga - Itarantim - Itiuba - Itororó - Itubera - Jaguaripe - João Dourado - Jussara - Jussarí - Jussiape - Lamarão - Lapão - Lençóis - Macarani - Maiquinique - Malhada - Maraú - Marcionílio Souza - Mascote - Matina - Monte Santo - Morpará - Morro do Chapeu - Mucugê - Mulungu do Morro - Muquém de São Francisco - Nilo Peçanha - Nordestina - Nova Canaã - Nova Redenção - Novo Horizonte - Oliveira dos Brejinhos - Palmeiras - Paratinga - Pau Brasil - Piatã - Piraí do Norte - Potiraguá - Presidente Dutra - Presidente Tancredo Neves - Queimadas - Quijingue - Retirolândia - Riacho de Santana - Rio de Contas - Santa Cruz da Vitoria - Santa Luzia - Santa luz - São Domingos - São Gabriel - São José da Vitória - Seabra - Serra do Ramalho - Serrinha - Sitio do Mato - Souto Soares - Taperoá - Teofilândia - Teolândia - Tucano - Ubaitaba - Uibaí - Una - Uruçuca - Utinga - Valença - Valente - Wagner - Wenceslau Guimaraes - Xique-Xique. -

Instituto De Meio Ambiente E Recursos Hídricos
Instituto de Meio Ambiente e Recursos Hídricos Diretoria de Recursos Hídricos e Monitoramento Ambiental Coordenação de Estudos de Clima e Projetos Especiais Precipitação Diária no Estado da Bahia - 19 de maio de 2020 - Ocorrência de chuvas nos postos monitorados pelo INEMA no período das 9h01 do dia 18/05/2020 às 9h00 do dia 19/05/2020 em mm. Chapada Diamantina Baixa Grande - PG-PR-08 7,60 Cafarnaum - 290530501C 6,20 Iaçu - ANA 7,60 Iaçu (João Amaro) - PG-PR- Iacú (Lagoa Bonita) - PG-PR- 2,80 4,00 Iaçu - PG-CL-05 1,60 16 15 Livramento de Nossa Senhora - Lençóis - 83242 4,00 Lençóis - A425 4,20 0,20 291950401U Marcionílio Souza (Machado 2,00 Morro do Chapéu - 83184 2,40 Mucugê (Guiné) - PG-PR-24 0,40 Portela) - PG- Mundo Novo (Ibiaporã) - PG- 19,80 Piatã - A430 7,60 Rio de Contas - 292670701C 1,20 CL-01 Seabra - PG-CL-09 0,20 Utinga - 293280401C 10,40 Nordeste Caém - IT-PR-35 2,20 Capim Grosso - IT-CL-03 0,60 Conde - A431 35,40 Crisópolis - IT-PR-08 4,20 Euclides da Cunha - A442 1,80 Heliópolis - 291185701C 1,60 Jacobina (Feliz Tomaz) - Inhambupe - RN-CL-10 2,60 Jacobina - A440 2,20 0,60 291750802A Jacobina (Itapeipu) - IT-PR- Nova Soure (Estação 1,20 Jeremoabo - A450 0,20 0,20 34 Experimental) - IT-PR-10 Quijingue (BR 116 E. Cunha / Pedro Alexandre - VB-PR-01 0,80 Ponto Novo 0,40 0,40 Tucano) - IT-PR- Ribeira do Amparo (Boa Ribeira do Pombal (Banzaê) - IT- Ribeira do Amparo - A458 1,00 2,60 1,00 Hora) - IT-PR-13 PR-01 Senhor do Bonfim (Carrapichel) Saúde - IT-CL-04 0,20 Senhor do Bonfim - A428 0,40 0,80 - IT-PR-37 Tucano (Caldas do Jorro)