Doctoral Thesis Hofstrom 2013
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Affibody Molecules for Epidermal Growth Factor Receptor Targeting
Journal of Nuclear Medicine, published on January 21, 2009 as doi:10.2967/jnumed.108.055525 Affibody Molecules for Epidermal Growth Factor Receptor Targeting In Vivo: Aspects of Dimerization and Labeling Chemistry Vladimir Tolmachev1-3, Mikaela Friedman4, Mattias Sandstrom¨ 5, Tove L.J. Eriksson2, Daniel Rosik2, Monika Hodik1, Stefan Sta˚hl4, Fredrik Y. Frejd1,2, and Anna Orlova1,2 1Unit of Biomedical Radiation Sciences, Rudbeck Laboratory, Uppsala University, Uppsala, Sweden; 2Affibody AB, Bromma, Sweden; 3Department of Medical Sciences, Nuclear Medicine, Uppsala University, Uppsala, Sweden; 4Division of Molecular Biotechnology, School of Biotechnology, Royal Institute of Technology, Stockholm, Sweden; and 5Section of Hospital Physics, Department of Oncology, Uppsala University Hospital, Uppsala, Sweden Noninvasive detection of epidermal growth factor receptor (EGFR) expression in malignant tumors by radionuclide molecu- he epidermal growth factor receptor (EGFR; other lar imaging may provide diagnostic information influencing pa- T designations are HER1 and ErbB-1) is a transmembrane tient management. The aim of this study was to evaluate a tyrosine kinase receptor that regulates cell proliferation, novel EGFR-targeting protein, the ZEGFR:1907 Affibody molecule, for radionuclide imaging of EGFR expression, to determine a motility, and suppression of apoptosis (1). Overexpression suitable tracer format (dimer or monomer) and optimal label. of EGFR is documented in several malignant tumors, such Methods: An EGFR-specific Affibody molecule, ZEGFR:1907, as carcinomas of the breast, urinary bladder, and lung, and 111 and its dimeric form, (ZEGFR:1907)2, were labeled with In using is associated with poor prognosis (2). A high level of EGFR 125 benzyl-diethylenetriaminepentaacetic acid and with I using expression could provide malignant cells with an advantage p-iodobenzoate. -

Monoclonal Antibody: a New Treatment Strategy Against Multiple Myeloma
antibodies Review Monoclonal Antibody: A New Treatment Strategy against Multiple Myeloma Shih-Feng Cho 1,2,3, Liang Lin 3, Lijie Xing 3,4, Tengteng Yu 3, Kenneth Wen 3, Kenneth C. Anderson 3 and Yu-Tzu Tai 3,* 1 Division of Hematology & Oncology, Department of Internal Medicine, Kaohsiung Medical University Hospital, Kaohsiung Medical University, Kaohsiung 807, Taiwan; [email protected] 2 Faculty of Medicine, College of Medicine, Kaohsiung Medical University, Kaohsiung 807, Taiwan 3 LeBow Institute for Myeloma Therapeutics and Jerome Lipper Multiple Myeloma Center, Dana-Farber Cancer Institute, Harvard Medical School, Boston, MA 02215, USA; [email protected] (L.L.); [email protected] (L.X.); [email protected] (T.Y.); [email protected] (K.W.); [email protected] (K.C.A.) 4 Department of Hematology, Shandong Provincial Hospital Affiliated to Shandong University, No. 324, Jingwu Road, Jinan 250021, China * Correspondence: [email protected]; Tel.: +1-617-632-3875; Fax: +1-617-632-2140 Received: 20 October 2017; Accepted: 10 November 2017; Published: 14 November 2017 Abstract: 2015 was a groundbreaking year for the multiple myeloma community partly due to the breakthrough approval of the first two monoclonal antibodies in the treatment for patients with relapsed and refractory disease. Despite early disappointments, monoclonal antibodies targeting CD38 (daratumumab) and signaling lymphocytic activation molecule F7 (SLAMF7) (elotuzumab) have become available for patients with multiple myeloma in the same year. Specifically, phase 3 clinical trials of combination therapies incorporating daratumumab or elotuzumab indicate both efficacy and a very favorable toxicity profile. These therapeutic monoclonal antibodies for multiple myeloma can kill target cells via antibody-dependent cell-mediated cytotoxicity, complement-dependent cytotoxicity, and antibody-dependent phagocytosis, as well as by direct blockade of signaling cascades. -

Opportunities for Conformation-Selective Antibodies in Amyloid-Related Diseases
Antibodies 2015, 4, 170-196; doi:10.3390/antib4030170 OPEN ACCESS antibodies ISSN 2073-4468 www.mdpi.com/journal/antibodies Review Opportunities for Conformation-Selective Antibodies in Amyloid-Related Diseases Marta Westwood * and Alastair D. G. Lawson Structural Biology, UCB, 216 Bath Road, Slough, SL1 3WE UK; E-Mail: [email protected]. * Author to whom correspondence should be addressed; E-Mail: [email protected]; Tel.: +44-1-753-534-655 (ext.7749); Fax: +44-1-753-536-632. Academic Editor: Dimiter S. Dimitrov Received: 13 May 2015 / Accepted: 9 July 2015 / Published: 15 July 2015 Abstract: Assembly of misfolded proteins into fibrillar deposits is a common feature of many neurodegenerative diseases. Developing effective therapies to these complex, and not yet fully understood diseases is currently one of the greatest medical challenges facing society. Slow and initially asymptomatic onset of neurodegenerative disorders requires profound understanding of the processes occurring at early stages of the disease including identification and structural characterisation of initial toxic species underlying neurodegeneration. In this review, we chart the latest progress made towards understanding the multifactorial process leading to amyloid formation and highlight efforts made in the development of therapeutic antibodies for the treatment of amyloid-based disorders. The specificity and selectivity of conformational antibodies make them attractive research probes to differentiate between transient states preceding formation of mature fibrils and enable strategies for potential therapeutic intervention to be considered. Keywords: antibody; amyloids; conformation; prion; Alzheimer’s; Parkinson’s; fibrils, tau; Huntingtin; protein misfolding 1. Introduction Correct protein folding is crucial for maintaining healthy biological functions. -

Rapid Selection of Specific MAP Kinase-Binders from Designed Ankyrin Repeat Protein Libraries
Protein Engineering, Design & Selection vol. 19 no. 5 pp. 219–229, 2006 Published online March 21, 2006 doi:10.1093/protein/gzl004 Rapid selection of specific MAP kinase-binders from designed ankyrin repeat protein libraries Patrick Amstutz1,4,5, Holger Koch1,4, H. Kaspar Binz1, form in this milieu. For this reason, antibodies were Stefan A. Deuber2 and Andreas Plu¨ckthun1,3 engineered for higher intracellular stability with some success 1 et al ¨ ¨ Biochemisches Institut der Universita¨tZu¨rich, Winterthurerstrasse 190, (Proba ., 1998; Worn and Pluckthun, 1998; Desiderio CH-8057 Zu¨rich, Switzerland and 2Institut fu¨r Medizinische Virologie der et al., 2001; Visintin et al., 2002), but the number of selected Universita¨tZu¨rich, Gloriastrasse 30, Zu¨rich, Switzerland and active binders is usually limited (Tanaka and Rabbitts, 3To whom correspondence should be addressed. 2003; Koch et al., 2006). These results can be explained by E-mail: [email protected] the selection pressure, which not only places demands on 4These authors contributed equally to this work. 5 specific binding but also requires stability under reducing Present address: Molecular Partners AG, c/o Universita¨tZu¨rich, conditions. Winterthurerstrasse 190, CH-8057 Zu¨rich, Switzerland To overcome these limitations of immunoglobulin We describe here the rapid selection of specific MAP-kinase domains, a variety of novel scaffolds for the generation of binders from a combinatorial library of designed ankyrin antibody-like binding molecules, some of them possessing repeat proteins (DARPins). A combined in vitro/in vivo selec- very favorable biophysical properties, has been developed tion approach, based on ribosome display and the protein (Nygren and Skerra, 2004; Binz et al., 2005). -

Blincyto Pi Hcp English.Pdf
HIGHLIGHTS OF PRESCRIBING INFORMATION These highlights do not include all the information needed to use BLINCYTO® safely and effectively. See full prescribing information for BLINCYTO. - Premedicate with dexamethasone. (2.2) BLINCYTO® (blinatumomab) for injection, for intravenous use • Refer to Full Prescribing Information for important preparation and Initial U.S. Approval: 2014 administration information. (2.4, 2.5, 2.6) • Administer as a continuous intravenous infusion at a constant flow rate using an infusion pump. (2.5, 2.6) WARNING: CYTOKINE RELEASE SYNDROME and - See Section 2.5 for infusion over 24 hours or 48 hours. NEUROLOGICAL TOXICITIES - See Section 2.6 for infusion over 7 days using Bacteriostatic 0.9% See full prescribing information for complete boxed warning. Sodium Chloride Injection, USP (containing 0.9% benzyl alcohol). This option is not recommended for patients weighing less than 22 kg. • Cytokine Release Syndrome (CRS), which may be life-threatening or fatal, occurred in patients receiving BLINCYTO. Interrupt or ---------------------DOSAGE FORMS AND STRENGTHS---------------------- discontinue BLINCYTO and treat with corticosteroids as For injection: 35 mcg of lyophilized powder in a single-dose vial for recommended. (2.3, 5.1) reconstitution. (3) • Neurological toxicities, which may be severe, life-threatening, or fatal, occurred in patients receiving BLINCYTO. Interrupt or discontinue -------------------------------CONTRAINDICATIONS------------------------------ BLINCYTO as recommended. (2.3, 5.2) Known hypersensitivity -

The Potential of Adoptive Transfer of Γ9δ2 T Cells to Enhance
www.nature.com/scientificreports OPEN The potential of adoptive transfer of γ9δ2 T cells to enhance blinatumomab’s antitumor activity against B‑cell malignancy Yun‑Hsiang Chen1, Yun Wang2, Cheng‑Hao Liao3 & Shu‑Ching Hsu4,5,6,7,8,9* Blinatumomab, a bispecifc T cell engager (BiTE) antibody targeting CD19 and CD3ε, can redirect T cells toward CD19‑positive tumor cells and has been approved to treat relapsed/refractory B‑cell acute lymphoblastic leukemia (R/R B‑ALL). However, chemotherapeutic regimens can severely reduce T cells’ number and cytotoxic function, leading to an inadequate response to blinatumomab treatment in patients. In addition, it was reported that a substantial portion of R/R B‑ALL patients failing blinatumomab treatment had the extramedullary disease, indicating the poor ability of blinatumomab in treating extramedullary disease. In this study, we investigated whether the adoptive transfer of ex vivo expanded γ9δ2 T cells could act as the efector of blinatumomab to enhance blinatumomab’s antitumor activity against B‑cell malignancies in vivo. Repeated infusion of blinatumomab and human γ9δ2 T cells led to more prolonged survival than that of blinatumomab or human γ9δ2 T cells alone in the mice xenografted with Raji cells. Furthermore, adoptive transfer of γ9δ2 T cells reduced tumor mass outside the bone marrow, indicating the potential of γ9δ2 T cells to eradicate the extramedullary disease. Our results suggest that the addition of γ9δ2 T cells to the blinatumomab treatment regimens could be an efective approach to enhancing blinatumomab’s therapeutic efcacy. The concept of this strategy may also be applied to other antigen‑specifc BiTE therapies for other malignancies. -

High Risk Therapy Made Easy: Supporting High Risk Patients Through Complex Therapy
8/21/2018 High Risk Therapy Made Easy: Supporting high risk patients through complex therapy Lori Ranney, MSN, APRN, CPNP, CPHON Mylynda Livingston, MSN, APRN, AC PC-PNP, CPON Teresa Herriage, DNP, APRN, CPNP, CPHON Children’s Minnesota Disclaimers and Confidentiality Protections Children’s Minnesota makes no representations or warranties about the accuracy, reliability, or completeness of the content. Content is provided “as is” and is for informational use only. It is not a substitute for professional medical advice, diagnosis, or treatment. Children’s disclaims all warranties, express or implied, statutory or otherwise, including without limitation the implied warranties of merchantability, non-infringement of third parties’ rights, and fitness for a particular purpose. This content was developed for use in Children’s patient care environment and may not be suitable for use in other patient care environments. Children’s does not endorse, certify, or assess third parties’ competency. You hold all responsibility for your use or nonuse of the content. Children’s shall not be liable for claims, losses, or damages arising from or related to any use or misuse of the content. This content and its related discussions are privileged and confidential under Minnesota’s peer review statute (Minn. Stat. § 145.61 et. seq.). Do not disclose unless appropriately authorized. Notwithstanding the foregoing, content may be subject to copyright or trademark law; use of such information requires Children’s permission. This content may include patient protected health information. You agree to comply with all applicable state and federal laws protecting patient privacy and security including the Minnesota Health Records Act and the Health Insurance Portability and Accountability Act and its implementing regulations as amended from time to time. -

Nanobodies Right in the Middle: Intrabodies As Toolbox to Visualize and Modulate Antigens in the Living Cell
biomolecules Review Nanobodies Right in the Middle: Intrabodies as Toolbox to Visualize and Modulate Antigens in the Living Cell Teresa R. Wagner 1,2 and Ulrich Rothbauer 1,2,* 1 Pharmaceutical Biotechnology, Eberhard Karls University Tuebingen, 72076 Tuebingen, Germany; [email protected] 2 Natural and Medical Sciences Institute, University of Tuebingen, 72770 Reutlingen, Germany * Correspondence: [email protected]; Tel.: +49-7121-5153-0415; Fax: +49-7121-5153-0816 Received: 30 November 2020; Accepted: 18 December 2020; Published: 21 December 2020 Abstract: In biomedical research, there is an ongoing demand for new technologies to elucidate disease mechanisms and develop novel therapeutics. This requires comprehensive understanding of cellular processes and their pathophysiology based on reliable information on abundance, localization, post-translational modifications and dynamic interactions of cellular components. Traceable intracellular binding molecules provide new opportunities for real-time cellular diagnostics. Most prominently, intrabodies derived from antibody fragments of heavy-chain only antibodies of camelids (nanobodies) have emerged as highly versatile and attractive probes to study and manipulate antigens within the context of living cells. In this review, we provide an overview on the selection, delivery and usage of intrabodies to visualize and monitor cellular antigens in living cells and organisms. Additionally, we summarize recent advances in the development of intrabodies as cellular biosensors and their application to manipulate disease-related cellular processes. Finally, we highlight switchable intrabodies, which open entirely new possibilities for real-time cell-based diagnostics including live-cell imaging, target validation and generation of precisely controllable binding reagents for future therapeutic applications. Keywords: nanobody; intrabody; phage display; live-cell imaging; biosensors; target validation 1. -
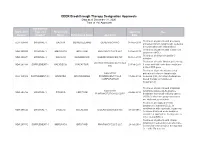
CDER Breakthrough Therapy Designation Approvals Data As of December 31, 2020 Total of 190 Approvals
CDER Breakthrough Therapy Designation Approvals Data as of December 31, 2020 Total of 190 Approvals Submission Application Type and Proprietary Approval Use Number Number Name Established Name Applicant Date Treatment of patients with previously BLA 125486 ORIGINAL-1 GAZYVA OBINUTUZUMAB GENENTECH INC 01-Nov-2013 untreated chronic lymphocytic leukemia in combination with chlorambucil Treatment of patients with mantle cell NDA 205552 ORIGINAL-1 IMBRUVICA IBRUTINIB PHARMACYCLICS LLC 13-Nov-2013 lymphoma (MCL) Treatment of chronic hepatitis C NDA 204671 ORIGINAL-1 SOVALDI SOFOSBUVIR GILEAD SCIENCES INC 06-Dec-2013 infection Treatment of cystic fibrosis patients age VERTEX PHARMACEUTICALS NDA 203188 SUPPLEMENT-4 KALYDECO IVACAFTOR 21-Feb-2014 6 years and older who have mutations INC in the CFTR gene Treatment of previously untreated NOVARTIS patients with chronic lymphocytic BLA 125326 SUPPLEMENT-60 ARZERRA OFATUMUMAB PHARMACEUTICALS 17-Apr-2014 leukemia (CLL) for whom fludarabine- CORPORATION based therapy is considered inappropriate Treatment of patients with anaplastic NOVARTIS lymphoma kinase (ALK)-positive NDA 205755 ORIGINAL-1 ZYKADIA CERITINIB 29-Apr-2014 PHARMACEUTICALS CORP metastatic non-small cell lung cancer (NSCLC) who have progressed on or are intolerant to crizotinib Treatment of relapsed chronic lymphocytic leukemia (CLL), in combination with rituximab, in patients NDA 206545 ORIGINAL-1 ZYDELIG IDELALISIB GILEAD SCIENCES INC 23-Jul-2014 for whom rituximab alone would be considered appropriate therapy due to other co-morbidities -

Designer Oncolytic Adenovirus: Coming of Age
Preprints (www.preprints.org) | NOT PEER-REVIEWED | Posted: 21 May 2018 doi:10.20944/preprints201805.0273.v1 Designer Oncolytic Adenovirus: Coming of Age Alexander T. Baker1, Carmen Aguirre-Hernandez2, Gunnel Hallden2, Alan L. Parker1* 1 Division of Cancer and Genetics, Cardiff University School of Medicine, Cardiff, CF14 4XN, United Kingdom 2 Centre for Molecular Oncology, Barts Cancer Institute, Queen Mary University of London, EC1M 6BQ, United Kingdom *Corresponding author Dr. Alan L. Parker Division of Cancer and Genetics Henry Wellcome Building Cardiff University School of Medicine Heath Park Cardiff CF14 4XN Email: [email protected] Keywords: adenovirus; oncolytic; targeting; virotherapy; cancer; αvβ6 integrin; immunotherapy; tropism 1 © 2018 by the author(s). Distributed under a Creative Commons CC BY license. Preprints (www.preprints.org) | NOT PEER-REVIEWED | Posted: 21 May 2018 doi:10.20944/preprints201805.0273.v1 Contents 1. Introduction: ................................................................................................................................... 4 2. Replication-selective adenoviruses ..................................................................................................... 6 2.1 Combination of oncolytic adenoviruses with chemotherapy ..................................................... 11 3. Oncolytic immunotherapy ................................................................................................................ 12 4. Tropism modification strategies ...................................................................................................... -

5.01.582 Antibody-Drug Conjugates
MEDICAL POLICY – 5.01.582 Antibody-Drug Conjugates Effective Date: June 1, 2021 RELATED MEDICAL POLICIES: Last Revised: May 11, 2021 None Replaces: N/A Select a hyperlink below to be directed to that section. POLICY CRITERIA | CODING | RELATED INFORMATION EVIDENCE REVIEW | REFERENCES | HISTORY ∞ Clicking this icon returns you to the hyperlinks menu above. Introduction An antibody is a blood protein. When the immune system detects an unhealthy cell, antibodies link to a molecule, known as an antigen, on the unhealthy cell. Monoclonal antibodies are produced in a laboratory. They are made to link to antigens usually found in high numbers on cancer cells. Antibody-drug conjugates combine monoclonal antibodies with certain chemotherapy drugs. The monoclonal antibodies find the cancer cells and the chemotherapy drug is released directly into those cells. The goal with this treatment is to target only cancer cells and spare nearby healthy cells. This policy describes when specific antibody-drug conjugates may be considered medically necessary. Note: The Introduction section is for your general knowledge and is not to be taken as policy coverage criteria. The rest of the policy uses specific words and concepts familiar to medical professionals. It is intended for providers. A provider can be a person, such as a doctor, nurse, psychologist, or dentist. A provider also can be a place where medical care is given, like a hospital, clinic, or lab. This policy informs them about when a service may be covered. Policy Coverage Criteria Drug Medical -

Article in Press
G Model MIMM-4561; No. of Pages 12 ARTICLE IN PRESS Molecular Immunology xxx (2015) xxx–xxx Contents lists available at ScienceDirect Molecular Immunology j ournal homepage: www.elsevier.com/locate/molimm Review Alternative molecular formats and therapeutic applications for ଝ bispecific antibodies ∗ Christoph Spiess, Qianting Zhai, Paul J. Carter Department of Antibody Engineering, Genentech Inc., 1 DNA Way, South San Francisco, CA 94080, USA a r t i c l e i n f o a b s t r a c t Article history: Bispecific antibodies are on the cusp of coming of age as therapeutics more than half a century after they Received 28 November 2014 ® were first described. Two bispecific antibodies, catumaxomab (Removab , anti-EpCAM × anti-CD3) and Received in revised form ® blinatumomab (Blincyto , anti-CD19 × anti-CD3) are approved for therapy, and >30 additional bispecific 30 December 2014 antibodies are currently in clinical development. Many of these investigational bispecific antibody drugs Accepted 2 January 2015 are designed to retarget T cells to kill tumor cells, whereas most others are intended to interact with two Available online xxx different disease mediators such as cell surface receptors, soluble ligands and other proteins. The modular architecture of antibodies has been exploited to create more than 60 different bispecific antibody formats. Keywords: These formats vary in many ways including their molecular weight, number of antigen-binding sites, Bispecific antibodies spatial relationship between different binding sites, valency for each antigen, ability to support secondary Antibody engineering Antibody therapeutics immune functions and pharmacokinetic half-life. These diverse formats provide great opportunity to tailor the design of bispecific antibodies to match the proposed mechanisms of action and the intended clinical application.