Noncoding Rnas and Their Annotation Using Metagenomics Algorithms Shubhra Sankar Ray1,2∗ and Sonam Maiti1
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Molecular Analysis of Small Rna and Small Protein Regulation of Escherichia Coli Stress Responses
MOLECULAR ANALYSIS OF SMALL RNA AND SMALL PROTEIN REGULATION OF ESCHERICHIA COLI STRESS RESPONSES BY CHELSEA R. LLOYD DISSERTATION Submitted in partial fulfillment of the requirements for the degree of Doctor of Philosophy in Microbiology in the Graduate College of the University of Illinois at Urbana-Champaign, 2018 Urbana, Illinois Doctoral Committee: Associate Professor Carin K. Vanderpool, Chair Professor John E. Cronan Professor William W. Metcalf Professor Peter A. B. Orlean Abstract Small RNA (sRNA) regulators control gene expression throughout all domains of life. In bacteria, they typically affect virulence, metabolism, and stress response genes posttranscriptionally through imperfect antisense pairing with their mRNAs. While most sRNAs are non-coding, a small number act as mRNAs themselves by encoding functional proteins. This study examines the regulatory and physiological effects of both a non-coding sRNA, DicF, and the protein product of a dual-function sRNA, SgrS in Eschericha coli. The sRNA SgrS encodes the small 43-amino acid protein SgrT. Both molecules are expressed during glucose-phosphate stress - a bacteriostatic condition in which phosphosugars accumulate in the cell either because of mutations in glycolysis or because of the transport of non-metabolizable glucose analogs such as αMG or 2DG. While both SgrT and SgrS base pairing can independently mitigate glucose-phosphate stress, they do so through distinct mechanisms. SgrS base pairing destabilizes the mRNA of the respective major and minor glucose transporters PtsG and ManXYZ, thereby inhibiting synthesis of additional glucose permeases and restricting further influx of non- metabolizable sugars. In this study we demonstrate that SgrT acts to specifically inhibit the transport activity of preexisting PtsG transporters, but does not affect ManXYZ. -

The Expression of NOD2, NLRP3 and NLRC5 and Renal Injury in Anti-Neutrophil Cytoplasmic Antibody-Associated Vasculitis
Wang et al. J Transl Med (2019) 17:197 https://doi.org/10.1186/s12967-019-1949-5 Journal of Translational Medicine RESEARCH Open Access The expression of NOD2, NLRP3 and NLRC5 and renal injury in anti-neutrophil cytoplasmic antibody-associated vasculitis Luo‑Yi Wang1,2,3, Xiao‑Jing Sun1,2,3, Min Chen1,2,3* and Ming‑Hui Zhao1,2,3,4 Abstract Background: Nucleotide‑binding oligomerization domain (NOD)‑like receptors (NLRs) are intracellular sensors of pathogens and molecules from damaged cells to regulate the infammatory response in the innate immune system. Emerging evidences suggested a potential role of NLRs in anti‑neutrophil cytoplasmic antibody (ANCA)‑associated vasculitis (AAV). This study aimed to investigate the expression of nucleotide‑binding oligomerization domain con‑ taining protein 2 (NOD2), NOD‑like receptor family pyrin domain containing 3 (NLRP3) and NOD‑like receptor family CARD domain containing 5 (NLRC5) in kidneys of AAV patients, and further explored their associations with clinical and pathological parameters. Methods: Thirty‑four AAV patients in active stage were recruited. Their renal specimens were processed with immu‑ nohistochemistry to assess the expression of three NLRs, and with double immunofuorescence to detect NLRs on intrinsic and infltrating cells. Analysis of gene expression was also adopted in cultured human podocytes. The associa‑ tions between expression of NLRs and clinicopathological parameters were analyzed. Results: The expression of NOD2, NLRP3 and NLRC5 was signifcantly higher in kidneys from AAV patients than those from normal controls, minimal change disease or class IV lupus nephritis. These NLRs co‑localized with podocytes and infltrating infammatory cells. -

Determinants of Target Prioritization and Regulatory Hierarchy for the Bacterial Small RNA Sgrs
Molecular Microbiology (2019) 112(4), 1199–1218 doi:10.1111/mmi.14355 First published online 6 August 2019 Determinants of target prioritization and regulatory hierarchy for the bacterial small RNA SgrS Maksym Bobrovskyy,1,† Muhammad S. Azam,1 of regulation of SgrS targets. The RNA chaperone Jane K. Frandsen,2,3,‡ Jichuan Zhang,4 Hfq uses distinct modes of binding to different SgrS Anustup Poddar,4 Xiangqian Ma,1 Tina M. Henkin,2 mRNA targets, which differentially influences posi- Taekjip Ha4,5 and Carin K. Vanderpool 1* tive and negative regulation. The RNA degradosome 1 Department of Microbiology, University of Illinois at plays a larger role in regulation of some SgrS targets Urbana-Champaign, 601 S. Goodwin Ave., Urbana, compared to others. Collectively, our results sug- IL 61801, USA. gest that sRNA selection of target mRNAs and regu- 2 Department of Microbiology and Center for RNA latory hierarchy are influenced by several molecular Biology, The Ohio State University, Columbus, features and that the combination of these features OH 43210, USA. precisely tunes the efficiency of regulation of multi- 3 Biochemistry Program, The Ohio State University, target sRNA regulons. Columbus, OH 43210, USA. 4 Department of Biophysics and Biophysical Chemistry, Johns Hopkins University, Baltimore, MD 21205, USA. Introduction 5 Howard Hughes Medical Institute, Baltimore, Bacteria live in diverse niches, often encountering rapidly MD 21205, USA. changing and stressful environments. Bacterial stress responses can mitigate the negative effects of stress on cell structure and function. Stress responses are usually coordinated by regulators, either RNAs or proteins, that Summary alter expression of a regulon comprised of multiple genes. -

Striking the Right Balance in Anti-Neutrophil Cytoplasmic Antibody-Associated Vasculitis
Striking the Right Balance in Anti-neutrophil Cytoplasmic Antibody-Associated Vasculitis This symposium took place on 4th June 2021, as part of the European Alliance of Associations for Rheumatology (EULAR) virtual congress Speakers: Benjamin Terrier,¹ Joanna Robson,² Bernhard Hellmich³ 1. University of Paris and Hôpital Cochin, France 2. University of the West of England and Bristol Royal Infirmary, UK 3. University of Tübingen, Germany Disclosure: Terrier has been an advisory board member and/or received consulting fees/ travel expenses from AstraZeneca, Chugai, Grifols, GlaxoSmithKline, Janssen, LFB, Octapharma, Roche, and Vifor Pharma. Robson has received speaker’s fees from Roche and Vifor Pharma; and research support from Vifor Pharma. Hellmich has been an investigator in clinical trials for Ab2Bio, AbbVie, AstraZeneca, Bristol- Myers Squibb, Chemocentrix, GlaxoSmithKline, InflaRx, Kiniksa, Nippon Kayaku, Novartis, Roche, and Sanofi. He has acted as a consultant, advisory board member, and/or lecturer for AbbVie, Bristol-Myers Squibb, Boehringer Ingelheim, Chugai, GlaxoSmithKline, InflaRx, Novartis, Pfizer, Roche, and Vifor Pharma. He is also a member of the Guideline Committees for European Alliance of Associations for Rheumatology (EULAR) and the German Society of Rheumatology (DGRh). Acknowledgements: Writing assistance was provided by Helen Boreham. Support: The publication of this article was funded by Vifor Pharma. The views and opinions expressed are those of the presenters. Content was reviewed by Vifor Pharma for medical accuracy. Citation: Rheumatol. 2021;8[1]:43-50. Meeting Summary Anti-neutrophil cytoplasmic antibody (ANCA)-associated vasculitis (AAV) causes irreversible short- and long-term damage to vital organs, particularly the kidneys and lungs. Current standard of care (SOC) for AAV, of which glucocorticoids (GC) are a lynchpin, has a number of important limitations: responses to therapy are variable, some patients fail to achieve and sustain remission, and treatment related adverse events (AE) are common. -

RNAIII of the Staphylococcus Aureus Agr System Activates Global Regulator Mgra by Stabilizing Mrna
Correction MICROBIOLOGY Correction for “RNAIII of the Staphylococcus aureus agr system (112:14036–14041; first published October 26, 2015; 10.1073/ activates global regulator MgrA by stabilizing mRNA,” by Ravi pnas.1509251112). Kr. Gupta, Thanh T. Luong, and Chia Y. Lee, which appeared The authors note that Fig. 2 appeared incorrectly. The cor- in issue 45, November 10, 2015, of Proc Natl Acad Sci USA rected figure and its legend appear below. A B 6 6 * * * 5 ** ** * 5 * 4 4 3 3 2 2 1 1 Half-life (min) 0 Half-life (min) 0 agr agr agr (11346) Newman P1 UTR P2 UTR P1 UTR P1 UTR P2 UTR P2 UTR (1844) (1845) +RNAIII +pML100 +RNAIII 5-nt mutant(12916) (A22) +pML100 (12501) (12502) P1 UTR P2 UTR 5-nt revertant Fig. 2. Stability of mgrA mRNA. (A) Stability in various chromosomal mutants. (B) RNAIII complementation of deletion mutations in mgrA UTR. mRNA stability expressed as half-life in minutes. The numbers in parentheses represent strain number. *P < 0.05, **P < 0.01 (unpaired two-tailed Student t test between Newman and each mutant, n = 3). www.pnas.org/cgi/doi/10.1073/pnas.1523895113 E7306 | PNAS | December 29, 2015 | vol. 112 | no. 52 www.pnas.org Downloaded by guest on September 26, 2021 RNAIII of the Staphylococcus aureus agr system activates global regulator MgrA by stabilizing mRNA Ravi Kr. Gupta, Thanh T. Luong, and Chia Y. Lee1 Department of Microbiology and Immunology, University of Arkansas for Medical Sciences, Little Rock, AR 72205 Edited by Richard P. Novick, New York University School of Medicine, New York, NY, and approved September 9, 2015 (received for review May 12, 2015) RNAIII, the effector of the agr quorum-sensing system, plays a key (13) to postulate that RNAIII must interact with one or more role in virulence gene regulation in Staphylococcus aureus, but how pleiotropic regulators. -
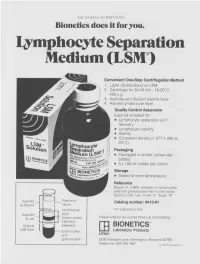
Lymphocyte Separation Medium (LSM
THE JOURNAL OF IMMUNOLOGY Bionetics does it for you. Lymphocyte Separation Medium (LSM wenient One-Step Centrifugation Method _ayer diluted blood on LSM. 2,entrifuge for 30-40 min., 18-20°C, ~.00 x g. ~,spirate and discard plasma layer -larvest lymphocyte layer. Quality Control Assurance Each lot is tested for: • Lymphocyte separation and recovery. • Lymphocyte viability. • Sterility. • Consistent density (1.077-1.080 at 20°C). Packaging • Packaged in amber, screw-cap bottles. • 5 x 100 ml bottles per carton. Storage. • Stored at room temperature. Reference Boyum, A. (1968): Isolation of mononuclear cells and granulocytes from human blood. Scand J. Clin. Lab. Invest. 21, Suppl. 97. Aspirate I IC[OI I IC~ LJI Catalog number: 8410-01 & discard serum Lymphocyte For Laboratory Use Aspirate layer & use (mononuclear Please write for our current Price List and Catalog. cells and Original platelets) ITi BIONETICS° LSM layer Erythrocytes Laboratory Products and Litton granulocytes 5516 Nicholson Lane, Kensington, Maryland 20795 Telephone: (301) 881-1557 1979 Litton Bionetics, tnc Get the most out of your high quality cytotoxic antibodies with LOW-TOX-M RABBIT COMPLEMENT LOW TOXICITY HIGH ACTIVITY Presentation: CL 3051 5 x 1 ml, lyophilized $30.00 When it comes to COMPLEMENT... come to CEDARLANE Direct orders or inquiries to: UNITED STATES: WORLDWIDE EXCEPT U.S. ,4 C~L CEDARLANE ACCURATE CHEMICAL & LABORATO RI ES SCIENTIFIC CORPORATION LIMITED 5516-8TH LINE, R.R. 2 28 TEC STREET, HICKSVtLLE, N.Y. 11801 HORNBY, ONTARIO, CANADA LOP 1E0 Telephone -

Genome-Wide Investigation of Micrornas and Their Targets in Response to Freezing Stress in Medicago Sativa L., Based on High-Throughput Sequencing
INVESTIGATION Genome-Wide Investigation of MicroRNAs and Their Targets in Response to Freezing Stress in Medicago sativa L., Based on High-Throughput Sequencing Yongjun Shu,1 Ying Liu, Wei Li, Lili Song, Jun Zhang, and Changhong Guo1 Key Laboratory of Molecular Cytogenetics and Genetic Breeding of Heilongjiang Province, College of Life Science and Technology, Harbin Normal University, 150025, China ABSTRACT Winter damage, especially in northern climates, is a major limitation of the utilization of perennial KEYWORDS forages such as alfalfa. Therefore, improving freezing tolerance is imperative in alfalfa genetic breeding. However, Medicago sativa freezing tolerance is a complex trait that is determined by many genes. To understand the complex regulation cold acclimation mechanisms of freezing tolerance in alfalfa, we performed small RNA sequencing analysis under cold (4°)and freezing stress freezing (28°) stress. The sequencing results revealed that 173 known, and 24 novel miRNAs were microRNA expressed, and that the expression of 35 miRNAs was affected by cold and/or freezing stress. Meanwhile, degradome 105 target genes cleaved by these miRNAs were characterized by degradome sequencing. These targets sequencing were associated with biological regulation, cellular processes, metabolic processes, and response to stress. Interestingly, most of them were characterized as transcription factors (TFs), including auxin response factors, SBP, NAC, AP2/ERF, and GRF, which play important roles in plant abiotic responses. In addition, important miRNAs and mRNAs involved in nodulation were also identified, for example, the relationship between miR169 and the TF CCAAT (also named as NF-YA/HAP2), which suggested that nodulation has an important function in freezing tolerance in alfalfa. -

Or Drought-Responsive Lncrnas in Cassava
www.nature.com/scientificreports OPEN Genome-wide identification and functional prediction of cold and/ or drought-responsive lncRNAs in Received: 06 December 2016 Accepted: 07 March 2017 cassava Published: 07 April 2017 Shuxia Li1,*, Xiang Yu2,3,*, Ning Lei1, Zhihao Cheng4, Pingjuan Zhao1, Yuke He2, Wenquan Wang1 & Ming Peng1 Cold and drought stresses seriously affect cassava (Manihot esculenta) plant growth and yield. Recently, long noncoding RNAs (lncRNAs) have emerged as key regulators of diverse cellular processes in mammals and plants. To date, no systematic screening of lncRNAs under abiotic stress and their regulatory roles in cassava has been reported. In this study, we present the first reference catalog of 682 high-confidence lncRNAs based on analysis of strand-specific RNA-seq data from cassava shoot apices and young leaves under cold, drought stress and control conditions. Among them, 16 lncRNAs were identified as putative target mimics of cassava known miRNAs. Additionally, by comparing with small RNA-seq data, we found 42 lncNATs and sense gene pairs can generate nat-siRNAs. We identified 318 lncRNAs responsive to cold and/or drought stress, which were typically co-expressed concordantly or discordantly with their neighboring genes. Trans-regulatory network analysis suggested that many lncRNAs were associated with hormone signal transduction, secondary metabolites biosynthesis, and sucrose metabolism pathway. The study provides an opportunity for future computational and experimental studies to uncover the functions of lncRNAs in cassava. Plants are sessile organisms and are constantly exposed to a wide range of environmental stresses during their life cycle. Cold and drought are the most severe abiotic stresses that seriously influence plant growth and develop- ment, and are major limiters of crop productivity worldwide1. -

Induction of Protein Citrullination and Auto-Antibodies Production In
www.nature.com/scientificreports OPEN Induction of protein citrullination and auto-antibodies production in murine exposed to nickel Received: 11 November 2015 Accepted: 21 December 2017 nanomaterials Published: xx xx xxxx Bashir M. Mohamed1,7, Noreen T. Boyle2,3, Anja Schinwald4, Bruno Murer5, Ronan Ward6, Omar K. Mahfoud1, Tatsiana Rakovich1, Kieran Crosbie-Staunton1, Steven G. Gray 7, Ken Donaldson4, Yuri Volkov1,2 & Adriele Prina-Mello 1,2 Citrullination, or the post-translational deimination of polypeptide-bound arginine, is involved in several pathological processes in the body, including autoimmunity and tumorigenesis. Recent studies have shown that nanomaterials can trigger protein citrullination, which might constitute a common pathogenic link to disease development. Here we demonstrated auto-antibody production in serum of nanomaterials-treated mice. Citrullination-associated phenomena and PAD levels were found to be elevated in nanomaterials -treated cell lines as well as in the spleen, kidneys and lymph nodes of mice, suggesting a systemic response to nanomaterials injection, and validated in human pleural and pericardial malignant mesothelioma (MM) samples. The observed systemic responses in mice exposed to nanomaterials support the evidence linking exposure to environmental factors with the development of autoimmunity responses and reinforces the need for comprehensive safety screening of nanomaterials. Furthermore, these nanomaterials induce pathological processes that mimic those observed in Pleural MM, and therefore require further investigations into their carcinogenicity. Citrullination is involved in several pathological processes in the body, including autoimmunity and tumor- igenesis. Citrullinated proteins are generated by a post-translational deimination or demethylimination of polypeptide-bound arginine by a family of Ca2+-dependent enzyme peptidylarginine deiminase (PAD)1. -

Computational Methods for the Identification and Characterization
Computational Methods for the Identification and Characterization of Non-Coding RNAs in Bacteria Dissertation der Mathematisch-Naturwissenschaftlichen Fakult¨at der Eberhard Karls Universit¨atT¨ubingen zur Erlangung des Grades eines Doktors der Naturwissenschaften (Dr. rer. nat.) vorgelegt von Alexander Herbig aus Altenkirchen T¨ubingen 2014 Tag der m¨undlichenQualifikation: 30.01.2015 Dekan: Prof. Dr. Wolfgang Rosenstiel 1. Berichterstatterin: PD Dr. Kay Nieselt 2. Berichterstatter: Prof. Dr. Daniel Huson 3. Berichterstatter: Prof. Dr. Rolf Backofen Zusammenfassung Forschungsergebnisse vergangener Jahre konnten zeigen wie komplex die Struktur und Regulation selbst bakterieller Transkriptome sein kann. Auch die wichtige Rolle nicht-kodierender RNAs (ncRNA), die nicht in Proteine translatiert werden, wird dabei immer deutlicher. Diese Molek¨uleerf¨ullen in der Zelle verschiedenste Aufgaben wie zum Beispiel die Regulation von Stoffwechselprozessen. Daher ist die Charakter- isierung der ncRNA-Gene eines Organismus immer mehr zu einem unverzichtbaren Teil von Systembiologie-Projekten geworden. Hierbei erlauben moderne Hochdurch- satzverfahren im Bereich der DNA- und RNA-Sequenzierung das im hohen Maße detaillierte Studium von Genomen und Transkriptomen. Die daraus resultierenden Daten m¨usseneiner vergleichenden Analyse unterzogen werden, um Variationen des Transkriptoms zwischen verschiedenen Organismen und Umweltbedingungen untersuchen zu k¨onnen.Hierf¨urwerden effiziente Computerprogramme ben¨otigt, die in der Lage sind genomische und transkriptomische Daten zu kombinieren und entsprechende Analysen automatisiert und reproduzierbar durchzuf¨uhren.Zu- dem m¨ussendiese Ans¨atzenicht-kodierende Elemente im genomischen Kontext lokalisieren und annotieren k¨onnen. In dieser Dissertation pr¨asentiere ich Computerprogramme zur L¨osungdieser Aufgaben. So wurde das Programm nocoRNAc entwickelt, welches ncRNAs in bakteriellen Genomen detektiert und diese bez¨uglich verschiedener Eigenschaften charakterisiert. -

The Ancestral Sgrs RNA Discriminates Horizontally Acquired Salmonella Mrnas Through a Single G-U Wobble Pair
The ancestral SgrS RNA discriminates horizontally acquired Salmonella mRNAs through a single G-U wobble pair Kai Papenforta, Dimitri Podkaminskia, Jay C. D. Hintonb, and Jörg Vogela,1 aRNA Biology Group, Institute for Molecular Infection Biology, University of Würzburg, D-97080 Würzburg, Germany; and bDepartment of Microbiology, Moyne Institute of Preventive Medicine, Trinity College, Dublin 2, Ireland AUTHOR SUMMARY Small noncoding RNAs A characteristic of HGT (sRNAs) constitute a vital group genes is that they are more likely of so-called posttranscriptional to undergo duplication than so- SgrS RNA regulators that shape the gene Core genome Horizontally acquired called core genes. Gene dupli- expression of eukaryotic and elements pathogenicity genes cation is a well-studied phe- prokaryotic organisms. In bac- nomenon accelerating evolu- teria, sRNAs generally act duplication tionary change in bacterial through base pairing to reduce pathogens (4). For example, the Hfq or increase the translation of ptsG/manXYZ mRNAs sopD mRNA sopD2 mRNA sopD gene has been duplicated target mRNAs into protein. target (G-C base-pair) pseudotarget (G-U base-pair) to generate sopD2 throughout Most of our current knowledge the S. enterica species, except in of sRNA numbers and functions the ancestral S. bongori (1). A Fig. P1. Posttranscriptional interaction between the core and stems from two species, Escher- horizontally acquired genome through Hfq and sRNAs. The SgrS sRNA bioinformatic comparison of the ichia coli and Salmonella enterica is encoded by the Salmonella core genome and conserved in many sopD and sopD2 sequences serovar Typhimurium. Both bacterial species, including E. coli and Salmonella. Aided by the RNA showed that the SgrS targeting organisms display a high degree chaperone Hfq, SgrS reduces the expression of mRNAs encoding for region is well-conserved be- of sequence conservation across sugar transport proteins (ptsG and manXYZ), both of which are core tween both genes; in other about three-quarters of the genomic elements. -

Evolution of the Small Family of Alternative Splicing Modulators Nuclear Speckle RNA-Binding Proteins in Plants
G C A T T A C G G C A T genes Article Evolution of the Small Family of Alternative Splicing Modulators Nuclear Speckle RNA-Binding Proteins in Plants Leandro Lucero 1, Jeremie Bazin 2, Johan Rodriguez Melo 3, Fernando Ibañez 3 , Martín D. Crespi 2,* and Federico Ariel 1,* 1 Instituto de Agrobiotecnología del Litoral, Universidad Nacional del Litoral, CONICET, FBCB, Centro Científico Tecnológico CONICET Santa Fe, Colectora Ruta Nacional No 168 km. 0, Paraje El Pozo, Santa Fe 3000, Argentina; [email protected] 2 CNRS, INRA, Institute of Plant Sciences Paris-Saclay IPS2, Universite Paris Sud, Universite Evry, Universite Paris-Diderot, Sorbonne Paris-Cite, Universite Paris-Saclay, 91405 Orsay, France; [email protected] 3 Instituto de Investigaciones Agrobiotecnológicas, CONICET, Universidad Nacional de Río Cuarto, Río Cuarto 5800, Argentina; [email protected] (J.R.M.); fi[email protected] (F.I.) * Correspondence: [email protected] (M.D.C.); [email protected] (F.A.); Tel./Fax: +54-342-4511-370 (ext. 5017) (F.A.) Received: 5 December 2019; Accepted: 30 January 2020; Published: 18 February 2020 Abstract: RNA-Binding Protein 1 (RBP1) was first identified as a protein partner of the long noncoding RNA (lncRNA) ENOD40 in Medicago truncatula, involved in symbiotic nodule development. RBP1 is localized in nuclear speckles and can be relocalized to the cytoplasm by the interaction with ENOD40. The two closest homologs to RBP1 in Arabidopsis thaliana were called Nuclear Speckle RNA-binding proteins (NSRs) and characterized as alternative splicing modulators of specific mRNAs.