Neurophysiological Correlates of Color Vision: a Model
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Diversity of Central Oxytocinergic Projections
Cell and Tissue Research (2019) 375:41–48 https://doi.org/10.1007/s00441-018-2960-5 REVIEW Diversity of central oxytocinergic projections Gustav F. Jirikowski1 Received: 21 September 2018 /Accepted: 6 November 2018 /Published online: 29 November 2018 # Springer-Verlag GmbH Germany, part of Springer Nature 2018 Abstract Localization and distribution of hypothalamic neurons expressing the nonapeptide oxytocin has been extensively studied. Their projections to the neurohypophyseal system release oxytocin into the systemic circulation thus controlling endocrine events associated with reproduction in males and females. Oxytocinergic neurons seem to be confined to the ventral hypothalamus in all mammals. Groups of such cells located outside the supraoptic and the paraventricular nuclei are summarized as Baccessory neurons.^ Although evolutionary probably associated with the classical magocellular nuclei, accessory oxytocin neurons seem to consist of rather heterogenous groups: Periventricular oxytocin neurons may gain contact to the third ventricle to secrete the peptide into the cerebrospinal fluid. Perivascular neurons may be involved in control of cerebral blood flow. They may also gain access to the portal circulation of the anterior pituitary lobe. Central projections of oxytocinergic neurons extend to portions of the limbic system, to the mesencephalon and to the brain stem. Such projections have been associated with control of behaviors, central stress response as well as motor and vegetative functions. Activity of the different oxytocinergic systems seems to be malleable to functional status, strongly influenced by systemic levels of steroid hormones. Keywords Hypothalamo neurohypophyseal system . Circumventricular organs . Liquor contacting neurons . Perivascular system . Limbic system Introduction been shown to occur in prostate, gonads, or skin, OTexpression in the brain seems to be confined to the hypothalamus. -

Quantitative Magnetic Resonance Imaging in Optic Neuritis
Quantitative magnetic resonance imaging in optic neuritis Simon James Hickman A thesis submitted to the University of London for the degree of Doctor of Philosophy 2004 NMR Research Unit Department of Neuroinflammation Institute of Neurology University College London Queen Square London WC1N3BG United Kingdom UMI Number: U602525 All rights reserved INFORMATION TO ALL USERS The quality of this reproduction is dependent upon the quality of the copy submitted. In the unlikely event that the author did not send a complete manuscript and there are missing pages, these will be noted. Also, if material had to be removed, a note will indicate the deletion. Dissertation Publishing UMI U602525 Published by ProQuest LLC 2014. Copyright in the Dissertation held by the Author. Microform Edition © ProQuest LLC. All rights reserved. This work is protected against unauthorized copying under Title 17, United States Code. ProQuest LLC 789 East Eisenhower Parkway P.O. Box 1346 Ann Arbor, Ml 48106-1346 i Abstract The major event in relapsing remitting multiple sclerosis (MS) is the acute relapse. The majority of patients with MS present with such an event. Most early relapses are followed by complete or near complete recovery, however partial recovery or significant disability can result from an individual relapse. Study of relapses in vivo is therefore desirable. Magnetic resonance imaging (MRI) is very sensitive at detecting MS lesions. Unfortunately, in the brain and spinal cord, it has been difficult to reliably identify the lesion that is responsible for the symptoms of an individual relapse and most new brain lesions which are apparent on MRI are clinically silent. -

Selective Cell Deathin Glaucoma
BritishJournal ofOphthalmology 1994; 78: 875-880 875 PERSPECTIVE Br J Ophthalmol: first published as 10.1136/bjo.78.11.875 on 1 November 1994. Downloaded from Selective cell death in glaucoma: does it really occur? J E Morgan Glaucoma is associated with retinal ganglion cell death and preparations2148 where, away from the fovea, cell size consequent deterioration in visual field. However, the asso- histograms indicated a reduction in the number ofcells at the ciation between these events is not straightforward since a upper end of the cell diameter range. A similar effect was substantial proportion ofthe ganglion cell population may be subsequently observed in tissue taken from the macular zone lost before visual field defects can be detected using conven- and examined in the transverse plane."9 It seemed reasonable tional perimetric methods.'2 Clinically, ganglion cell loss to assume that these changes in the axon and cell diameter manifests as optic disc cupping and defects in the nerve fibre distributions would be reflected as a change in the proportion layer3 before the onset ofvisual field changes. In recent years a ofthe types of remaining ganglion cells. number of studies have suggested that some ganglion cells The first aspect ofthese cell size distributions to consider is may have a greater susceptibility to the damaging effects of whether they result from a mechanism of cell death that is glaucoma and this idea has become central to a number of influenced by cell size. Taken on its own, this point is strategies designed to detect the earliest changes in this important since it has implications for the mechanism of cell disease. -
![Arxiv:1907.02116V1 [Q-Bio.NC] 3 Jul 2019](https://docslib.b-cdn.net/cover/4734/arxiv-1907-02116v1-q-bio-nc-3-jul-2019-1184734.webp)
Arxiv:1907.02116V1 [Q-Bio.NC] 3 Jul 2019
Multiplicative modulations in hue-selective cells enhance unique hue representation Paria Mehrani1, 2, *, Andrei Mouraviev1, 2, and John K. Tsotsos1, 2 1Department of Electrical Engineering and Computer Science, York University, Toronto, Canada 2The Center for Vision Research, York University, Toronto, Canada [email protected], [email protected], [email protected] There is still much to understand about the color processing mechanisms in the brain and the transformation from cone-opponent representations to perceptual hues. Moreover, it is unclear which areas(s) in the brain represent unique hues. We propose a hierarchical model inspired by the neuronal mechanisms in the brain for local hue representation, which reveals the contributions of each visual cortical area in hue representation. Local hue encoding is achieved through incrementally increasing processing nonlinearities beginning with cone input. Besides employing nonlinear rectifications, we propose multiplicative modulations as a form of nonlinearity. Our simulation results indicate that multiplicative modulations have significant contributions in encoding of hues along intermediate directions in the MacLeod-Boynton diagram and that model V4 neurons have the capacity to encode unique hues. Additionally, responses of our model neurons resemble those of biological color cells, suggesting that our model provides a novel formulation of the brain's color processing pathway. The color processing mechanisms in the primary visual Beyond LGN, studies on multiple regions in the ventral cortex and later processing stages are a target of debate stream show an increase in nonlinearity with respect to among color vision researchers. What is also unclear is the three cone types from LGN to higher brain areas [7] which brain area represents unique hues, those pure col- and also a shift of selectivity toward intermediate hues ors unmixed with other colors. -

Arginine Vasotocin Neuronal Phenotypes Among Congeneric Territorial and Shoaling Reef Butterflyfishes: Species, Sex and Reproductive Season Comparisons
Journal of Neuroendocrinology From Molecular to Translational Neurobiology Journal of Neuroendocrinology 20, 1382À1394 ORIGINAL ARTICLE ª 2008 The Authors. Journal Compilation ª 2008 Blackwell Publishing Ltd Arginine Vasotocin Neuronal Phenotypes among Congeneric Territorial and Shoaling Reef Butterflyfishes: Species, Sex and Reproductive Season Comparisons A. K. Dewan, K. P. Maruska and T. C. Tricas Department of Zoology, University of Hawai’i at Manoa, Honolulu and Hawai’i Institute of Marine Biology, Kaneohe, HI, USA. Journal of Arginine vasotocin (AVT) and the homologous arginine vasopressin (AVP) neuropeptides are involved in the control of aggression, spacing behaviour and mating systems in vertebrates, but Neuroendocrinology the function of AVT in the regulation of social behaviour among closely-related fish species needs further clarification. We used immunocytochemical techniques to test whether AVT neuro- nes show species, sex or seasonal differences in two sympatric butterflyfish sister species: the territorial monogamous multiband butterflyfish, Chaetodon multicinctus, and the shoaling polyg- amous milletseed butterflyfish, Chaetodon miliaris. The territorial species had larger AVT-immu- noreactive (-ir) somata within the preoptic area, and higher AVT fibre densities within but not limited to the ventral telencephalon, medial and dorsal nucleus of the dorsal telencephalon, torus semicircularis, and tectum compared to the shoaling nonterritorial species. Furthermore, AVT-ir somata size and number did not differ among sexes or spawning periods in the territorial species, and showed only limited variation within the shoaling species. The distinct difference in AVT neuronal characteristics among species is likely to be independent of body size differences, and the lack of sex and seasonal variability is consistent with their divergent but stable social and mating systems. -

Identifying Glaucomatous Vision Loss with Visual-Function–Specific
Identifying Glaucomatous Vision Loss with Visual-Function–Specific Perimetry in the Diagnostic Innovations in Glaucoma Study Pamela A. Sample, Felipe A. Medeiros, Lyne Racette, John P. Pascual, Catherine Boden, Linda M. Zangwill, Christopher Bowd, and Robert N. Weinreb PURPOSE. To compare the diagnostic results of four perimetric rameters at suggested criterion values provided good sensitiv- tests and to identify useful parameters from each for determin- ity and specificity. FDT showed the highest sensitivity overall, ing abnormality. with SAP performing better than in prior reports. Of note, the METHODS. One hundred eleven eyes with glaucomatous optic same area of the retina was identified as damaged in all tests. neuropathy (GON), 31 with progressive optic neuropathy (Invest Ophthalmol Vis Sci. 2006;47:3381–3389) DOI: (PGON) 53 with ocular hypertension, and 51 with no disease 10.1167/iovs.05-1546 were included (N ϭ 246). Visual field results were not used to classify the eyes. Short-wavelength automated perimetry ver the past several years, psychophysical tests of specific (SWAP), frequency-doubling technology perimetry (FDT), Ovisual functions have been used to measure visual perfor- high-pass resolution perimetry (HPRP), and standard auto- mance and to understand the underlying glaucomatous mated perimetry (SAP) were performed. Receiver operating changes in retinal ganglion cell function. Testing vision with characteristic (ROC) curves were used to compute the areas standard automated perimetry (SAP) is not selective for a par- under the curves (AUC) and sensitivity levels at given specific- ticular ganglion cell type. Any of the primary ganglion cell ities for a variety of abnormality criteria. The agreement among subtypes can respond to an achromatic incremental stimulus tests for abnormality, location, and extent of visual field deficit presented on an achromatic background. -

Representation of Perceptual Color Space in Macaque Posterior Inferior Temporal Cortex (The V4 Complex)
New Research Sensory and Motor Systems Representation of Perceptual Color Space in Macaque Posterior Inferior Temporal Cortex (the V4 Complex) †,Thorsten Hansen,3 and Bevil R. Conway1,2 ء,Katherine L. Hermann,1,2 ء,Kaitlin S. Bohon,1,2 DOI:http://dx.doi.org/10.1523/ENEURO.0039-16.2016 1Program in Neuroscience, Wellesley College, Wellesley, Massachusetts 02481, 2Department of Brain and Cognitive Sciences, Massachusetts Institute of Technology, Cambridge, Massachusetts 02139, and 3Department of Psychology, Justus Liebig University Giessen, 35396 Giessen, Germany Abstract The lateral geniculate nucleus is thought to represent color using two populations of cone-opponent neurons [L vs M; Svs(Lϩ M)], which establish the cardinal directions in color space (reddish vs cyan; lavender vs lime). How is this representation transformed to bring about color perception? Prior work implicates populations of glob cells in posterior inferior temporal cortex (PIT; the V4 complex), but the correspondence between the neural representation of color in PIT/V4 complex and the organization of perceptual color space is unclear. We compared color-tuning data for populations of glob cells and interglob cells to predictions obtained using models that varied in the color-tuning narrowness of the cells, and the color preference distribution across the populations. Glob cells were best accounted for by simulated neurons that have nonlinear (narrow) tuning and, as a population, represent a color space designed to be perceptually uniform (CIELUV). Multidimensional scaling and representational similarity analyses showed that the color space representations in both glob and interglob populations were correlated with the organization of CIELUV space, but glob cells showed a stronger correlation. -

Effects of Early Unilateral Blur on the Macaque's Visual System. II
The Journal of Neuroscience, May 1987, 7(5): 1327-l 339 Effects of Early Unilateral Blur on the Macaque’s Visual System. II. Anatomical Observations Anita E. Hendrickson,lB2 J. Anthony Movshon, 4 Howard M. Eggers,5 Martin S. Gizzi,4,a Ronald G. Boothe,3,b and Lynne Kiorpes3zc Departments of ‘Biological Structure, 20phthalmology, and 3Psychology, University of Washington, Seattle, Washington 98195; 4Department of Psychology, New York University, New York, New York 10003; and 5Harkness Eye Institute, College of Physicians and Surgeons, Columbia University, New York, New York 10036 We studied the effects of early unilateral blur on the ana- natal blur preferentially affects the parvocellular layers of tomical organization of the visual pathways in 8 macaque the LGN and layer 4Cfl of striate cortex, which are the por- monkeys. Blur was induced in one eye, beginning 2-14 d af- tions of the central visual system associated with the pro- ter birth, by 0.5% atropine twice a day. Atropiniration was cessing of information concerning fine spatial detail. These stopped at 6-8 months of age, and the animals were studied anatomical changes are consistent with the high spatial fre- for anatomy 3-24 months later. The retina and all other eye quency loss of vision demonstrated behaviorally and elec- tissues showed normal histology. In the dorsal lateral ge- trophysiologically in the atropine eye-driven visual system niculate nucleus (LGN), cells in parvocellular layers receiv- of these same animals. ing input from the atropine-treated eye were 9-32% smaller and were more lightly stained than those in layers innervated Amblyopia is a lossof visual function that is not due to optical by the untreated eye. -

The Distribution of Oxytocin and the Oxytocin Receptor in Rat Brain: Relation to Regions Active in Migraine
Warfvinge et al. The Journal of Headache and Pain (2020) 21:10 The Journal of Headache https://doi.org/10.1186/s10194-020-1079-8 and Pain RESEARCH ARTICLE Open Access The distribution of oxytocin and the oxytocin receptor in rat brain: relation to regions active in migraine Karin Warfvinge1,2* , Diana Krause2,3 and Lars Edvinsson1,2 Abstract Background: Recent work, both clinical and experimental, suggests that the hypothalamic hormone oxytocin (OT) and its receptor (OTR) may be involved in migraine pathophysiology. In order to better understand possible central actions of OT in migraine/headache pathogenesis, we mapped the distribution of OT and OTR in nerve cells and fibers in rat brain with a focus on areas related to migraine attacks and/or shown previously to contain calcitonin gene related peptide (CGRP), another neuropeptide involved in migraine. Methods: Distribution of OT and OTR in the adult, rat brain was qualitatively examined with immunohistochemistry using a series of well characterized specific antibodies. Results: As expected, OT was extensively localized in the cell somas of two hypothalamic nuclei, the supraoptic (SO or SON) and paraventricular nuclei (Pa or PVN). OT also was found in many other regions of the brain where it was localized mainly in nerve fibers. In contrast, OTR staining in the brain was mainly observed in cell somas with very little expression in fibers. The most distinct OTR expression was found in the hippocampus, the pons and the substantia nigra. In some regions of the brain (e.g. the amygdala and the hypothalamus), both OT and OTR were expressed (match). -

Perinatal Exposure to Organohalogen Pollutants Decreases Vasopressin
UC Riverside UC Riverside Previously Published Works Title Perinatal exposure to organohalogen pollutants decreases vasopressin content and its mRNA expression in magnocellular neuroendocrine cells activated by osmotic stress in adult rats. Permalink https://escholarship.org/uc/item/2093v8pq Journal Toxicology and applied pharmacology, 329 ISSN 0041-008X Authors Mucio-Ramírez, Samuel Sánchez-Islas, Eduardo Sánchez-Jaramillo, Edith et al. Publication Date 2017-08-01 DOI 10.1016/j.taap.2017.05.039 Peer reviewed eScholarship.org Powered by the California Digital Library University of California Toxicology and Applied Pharmacology 329 (2017) 173–189 Contents lists available at ScienceDirect Toxicology and Applied Pharmacology journal homepage: www.elsevier.com/locate/taap Perinatal exposure to organohalogen pollutants decreases vasopressin content and its mRNA expression in magnocellular neuroendocrine cells activated by osmotic stress in adult rats Samuel Mucio-Ramírez a,EduardoSánchez-Islasa, Edith Sánchez-Jaramillo b, Margarita Currás-Collazo c, Victor R. Juárez-González d, Mhar Y. Álvarez-González a, L.E. Orser c, Borin Hou c, Francisco Pellicer e, Prasada Rao S. Kodavanti f,MarthaLeón-Oleaa,⁎ a Departamento de Neuromorfología Funcional, Dirección de Investigaciones en Neurociencias, Instituto Nacional de Psiquiatría Ramón de la Fuente Muñiz, Calz. México Xochimilco No. 101, Col. San Lorenzo Huipulco, México D.F. C.P. 14370, México b Laboratorio de Neuroendocrinología Molecular, Dirección de Investigaciones en Neurociencias, Instituto Nacional de Psiquiatría Ramón de la Fuente Muñiz, Calz. México Xochimilco No. 101, Col. San Lorenzo Huipulco. México D.F. C.P. 14370, México c Department of Cell Biology and Neuroscience, University of California, Riverside, CA 92521, USA d Medicina Molecular y Bioprocesos, Instituto de Biotecnología, UNAM, Av. -
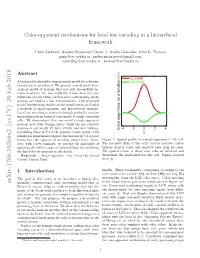
Color-Opponent Mechanisms for Local Hue Encoding in a Hierarchical Framework
Color-opponent mechanisms for local hue encoding in a hierarchical framework Paria Mehrani,∗ Andrei Mouraviev,∗ Oscar J. Avella Gonzalez, John K. Tsotsos [email protected], [email protected], [email protected] , [email protected] Abstract L cone A biologically plausible computational model for color rep- M cone resentation is introduced. We present a mechanistic hier- archical model of neurons that not only successfully en- codes local hue, but also explicitly reveals how the con- tributions of each visual cortical layer participating in the process can lead to a hue representation. Our proposed model benefits from studies on the visual cortex and builds a network of single-opponent and hue-selective neurons. Local hue encoding is achieved through gradually increas- ing nonlinearity in terms of cone inputs to single-opponent cells. We demonstrate that our model's single-opponent neurons have wide tuning curves, while the hue-selective neurons in our model V4 layer exhibit narrower tunings, Cone responses as a function of x -4 -2 0 2 4 resembling those in V4 of the primate visual system. Our x simulation experiments suggest that neurons in V4 or later layers have the capacity of encoding unique hues. More- Figure 1: Spatial profile of a single-opponent L+M- cell. over, with a few examples, we present the possibility of The receptive field of this cells receives positive contri- spanning the infinite space of physical hues by combining butions from L cones and negative ones from M cones. the hue-selective neurons in our model. The spatial extent of these cone cells are different and Keywords| Single-opponent, Hue, Hierarchy, Visual determines the mechanism for this cell. -

Multiplicative Modulations Enhance Diversity of Hue-Selective Cells Paria Mehrani✉, Andrei Mouraviev & John K
www.nature.com/scientificreports OPEN Multiplicative modulations enhance diversity of hue-selective cells Paria Mehrani✉, Andrei Mouraviev & John K. Tsotsos There is still much to understand about the brain’s colour processing mechanisms and the transformation from cone-opponent representations to perceptual hues. Moreover, it is unclear which area(s) in the brain represent unique hues. We propose a hierarchical model inspired by the neuronal mechanisms in the brain for local hue representation, which reveals the contributions of each visual cortical area in hue representation. Hue encoding is achieved through incrementally increasing processing nonlinearities beginning with cone input. Besides employing nonlinear rectifcations, we propose multiplicative modulations as a form of nonlinearity. Our simulation results indicate that multiplicative modulations have signifcant contributions in encoding of hues along intermediate directions in the MacLeod-Boynton diagram and that our model V2 neurons have the capacity to encode unique hues. Additionally, responses of our model neurons resemble those of biological colour cells, suggesting that our model provides a novel formulation of the brain’s colour processing pathway. Te colour processing mechanisms in the primary visual cortex and later processing stages are a target of debate among colour vision researchers. What is also unclear is which brain area represents unique hues, those pure colours unmixed with other colours. In spite of a lack of agreement on colour mechanisms in higher visual areas, human visual system studies confrm that colour encoding starts with three types of cones sensitive to long (L), medium (M) and short (S) wavelengths, forming the LMS colour space. Cones send opponent feedforward sig- nals to LGN cells with single-opponent receptive felds1–3.